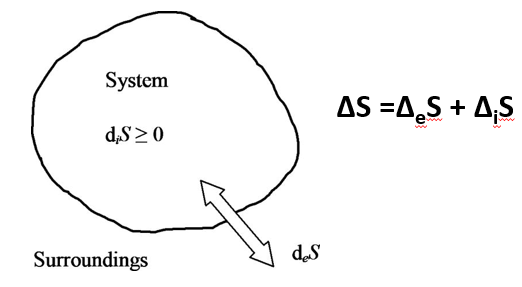
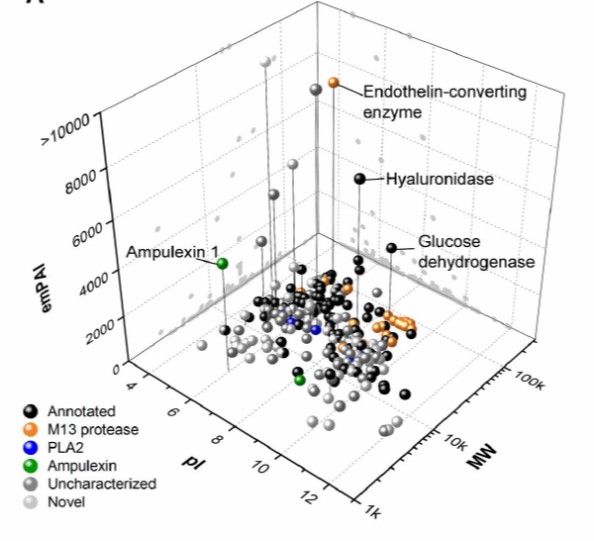
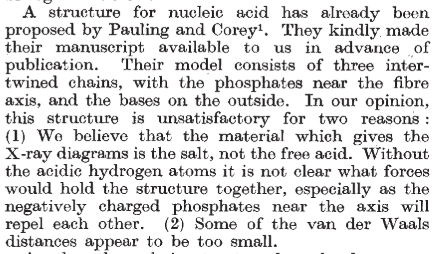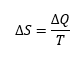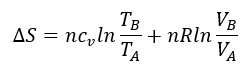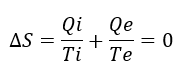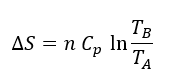Cambiando la perspectiva: la glucolisis como ciclo autocatalítico y la entropía de la vida.
escrito por C. Menor-Salvan | 3 octubre, 2023
C. Menor-Salván. Octubre 2023
«The best that most of us can hope to achieve in Science is simply to misunderstand at a deeper level»
Wolfgang Pauli
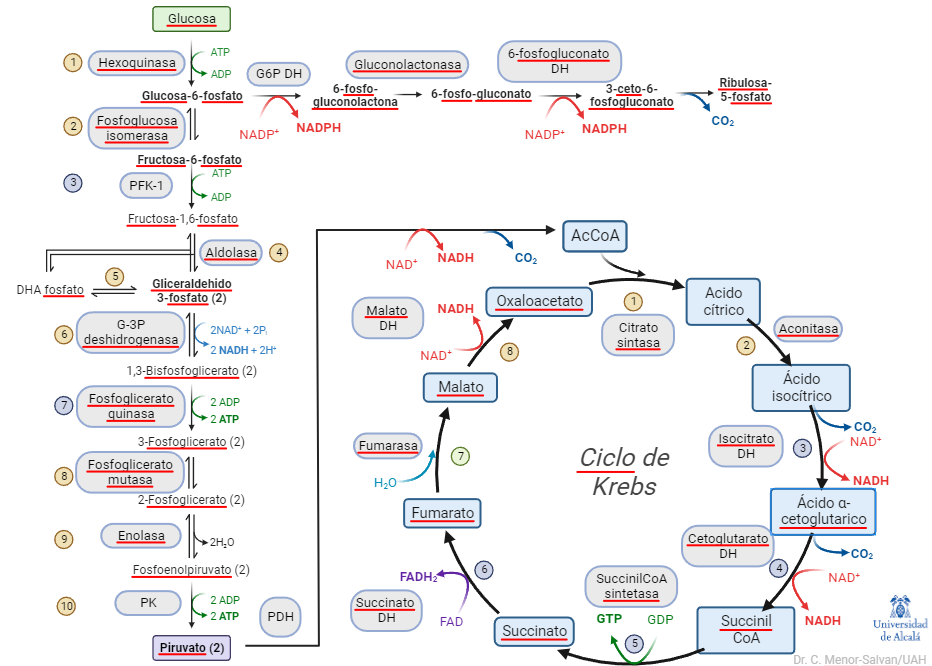
En los cursos introductorios y de bioquímica general, se suele explicar el metabolismo central como una complicada serie de reacciones químicas, que los estudiantes, en ocasiones, son casi forzados a memorizar. En esta exposición del metabolismo, la glucólisis se muestra como una ruta lineal, desde la glucosa al piruvato, que conecta con el ciclo de Krebs y con la ruta de las pentosas-fosfato.
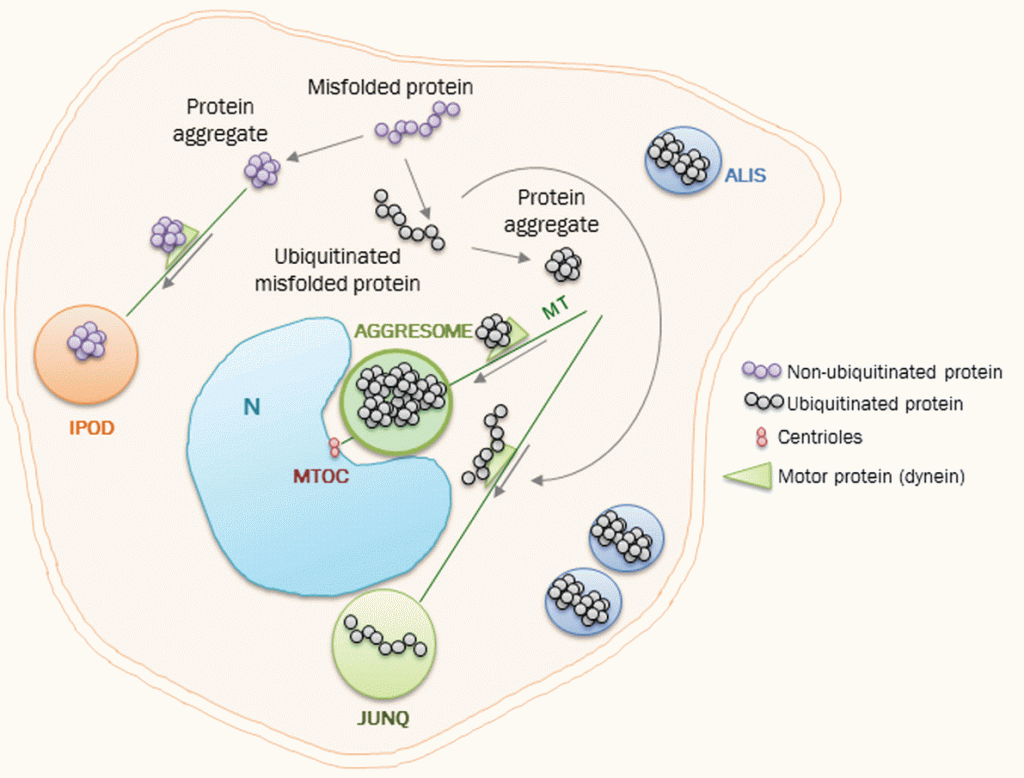
Glucolisis y ciclo de Krebs, junto con parte de la rama oxidativa del ciclo de las pentosas fosfato. En esta representación tradicional, la glucolisis aparece como ruta lineal.
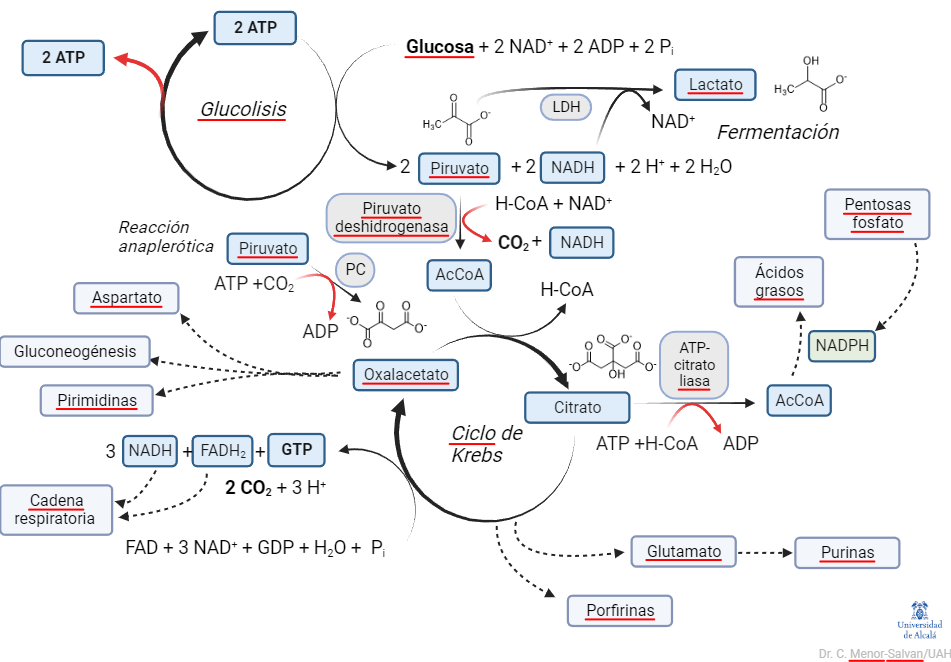
El problema de esta representación es que no da una visión sintética u holística de la glucolisis en el metabolismo. Desglosa sus reacciones y muestra su papel mecánico, pero ignora un aspecto esencial para la organización de un organismo vivo. El desglose de las reacciones, no obstante, ya nos da una pista: la glucolisis es una serie de reacciones reversibles e irreversibles. Y el relato de su auto-regulación nos muestra la importancia del control de sus pasos irreversibles. Si simplificamos la representación y la cambiamos ligeramente de perspectiva, podemos ver el metabolismo central así:
Vista muy simplificada del metabolismo. Las líneas punteadas indican rutas conectadas. Los componentes de la ruta glucolítica también tienen conexión con diversas rutas, pero no se muestran para no aumentar la complejidad de la imagen. Mas adelante hablaremos de la importancia de las ramas fermentativa y respiratoria, en relación con la variación del flujo de entropía en el cáncer.
¿Qué vemos en esta representación? Primero, resaltamos el papel del ciclo de Krebs: es la «M40 del metabolismo». Como una carretera de circunvalación en una ciudad muy poblada, distribuye el tráfico de carbono entre diversas rutas. También, impulsa la transformación de la energía del azúcar en hidrógeno, en pasos oxidativos claves. Este hidrógeno (en forma de cofactores NADH y FADH2), a su vez, mantendrá un gradiente, cuya disipación mueve la ATPasa, la dinamo del metabolismo.
Segundo, la glucolisis no aparece como una ruta lineal, sino como un ciclo de reacciones. Además, es un ciclo peculiar, en el que los productos del ciclo son, a su vez, reactivos. Una reacción, o ciclo de reacciones que representa una reacción global, en la que los productos son reactivos de la reacción se denomina reacción autocatalítica o ciclo autocatalítico.
Podemos resumir la glucolisis en una reacción global:
Glucosa + 2 NAD+ + 2 ADP + 2 ATP + 2 Pi –> 2 piruvato + 2 NADH + 4 ATP + 2 H+ + 2 H2O
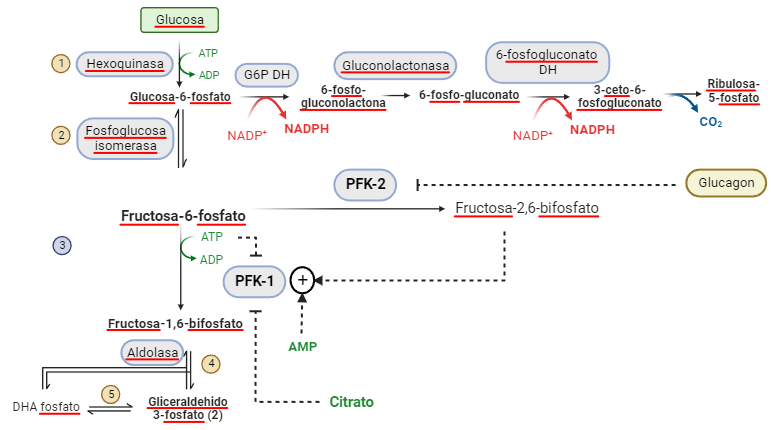
Los ciclos autocatalíticos tienen unas propiedades peculiares. Estas propiedades surgen como consecuencia de que estos sistemas se alejan del equilibrio. En el ciclo, gradualmente acelerado, se producen dos fenómenos superpuestos: las etapas formadas por reacciones irreversibles y que constituyen puntos de control, y los fenónenos no lineales de transporte de los componentes (difusión de ADP y ATP y disponibilidad de glucosa por ejemplo). La reacción pasa un umbral de inestabilidad tras el cual, surgen comportamientos complejos: oscilaciones periódicas, caos en la formación de los productos, ruptura de la homogeneidad, creación de ondas de diferente composición, cooperación molecular a nivel macroscópico y morfogénesis. La glucolisis y su regulación entonces se convierte en el control crucial de la formación de toda la estructura metabólica.
Control de la glucolisis por la PFK-1 y 2. La concentración de Fructosa-1,6-bifosfato controla la oscilación de la glucolisis, y el régimen caótico o periódico en la producción de ATP. Es interesante que la reacción de la PFK-1 es la que mayor entropía genera en la ruta.
La vida se sostiene sobre este alejamiento del equilibrio, y las células manifiestan varios ciclos autocatalíticos esenciales y sistemas alejados del equilibrio: la autoorganización de membranas impulsada por las propias membranas, o la síntesis de ácidos nucleicos, la señalización celular, los impulsos nerviosos… etc.
Estos comportamientos complejos alejados del equilibrio pueden dar lugar a la creación de estructuras organizadas, pero no siempre tiene que ser así. Cuando un sistema autocatalítico sobrepasa el umbral de inestabilidad, se bifurca en dos posibles estados: uno de los estados puede ser caótico, y el otro estado puede conducir a oscilaciones periódicas y cooperación molecular, y ambas pueden sucederse una a otra en sucesivas bifurcaciones.
Bioanálogos
Hay dos modelos de reacciones autocatalíticas que ilustran ambos comportamientos. El primero es la reacción de la formosa, una reacción autocatalítica de síntesis de azúcares a partir de formaldehído que entra en régimen caótico y conduce a una explosión combinatorial, resultando en una mezcla desorganizada de centenares de componentes. Podemos ilustrarlo con una analogía física: si acoplamos un micrófono y un altavoz, aunque la sala esté en silencio, las pequeñas perturbaciones que generan ruido en el altavoz, son amplificadas autocatalíticamente hasta producir un ruido ensordecedor.
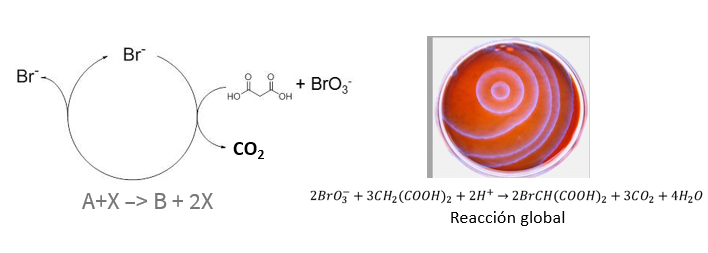
El segundo es una de las reacciones químicas más importantes del siglo XX: la reacción de Belousov-Zhabotinsky. Esta reacción es una ‘maqueta’ del metabolismo sostenida por una reacción autocatalítica. Hablamos de ella algo más en nuestra entrada sobre entropía.
Reacción de Belousov-Zhabotinsky. En realidad, la reacción tiene varias versiones, pero el mismo fundamento: un sistema de reacciones autocatalítico, que conecta una red de unas 40 a 80 reacciones químicas. Esta reacción es un bioanálogo, un sistema que funciona de modo similar a la vida y que nos ayuda a entenderla.
Esta reacción es una buena forma de visualizar qué ocurre cuando un sistema se aleja del equilibrio y se produce una ruptura de simetría. Esto es, cómo, desde una mezcla homogénea original, se produce espontáneamente una compartimentalización con diferentes especies y reacciones. En definitiva, cómo un sistema de reacciones impulsa que surjan estructuras organizadas. Estas estructuras se llaman patrones de Turing.
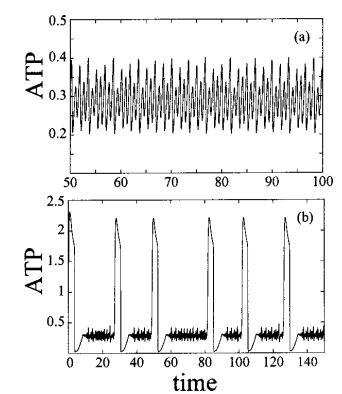
Al igual que en la reacción de Belousov, la glucolisis genera patrones de Turing en la concentración celular de ATP, que actúa como señal intracelular y controla toda la estructura. Un sistema caótico, como son estas reacciones, dependen en gran medida de las condiciones iniciales. Si se arranca una serie temporal con la glucolisis a dos concentraciones distintas de fructosa-1,6-bifosfato, el patrón de producción de ATP es muy diferente:
El incremento de la concentración inicial de F-1,6-BP cambia el patrón temporal de la glucolisis desde (a) oscilaciones caóticas a (b), picos de alta producción de ATP, seguidos de periodo de caos de pequeña amplitud. Figura de Baier, G., Müller, M. & Ørsnes, H. (2002) ‘Excitable Spatio-Temporal Chaos in a Model of Glycolysis’, The Journal of Physical Chemistry B, 106(12), pp. 3275–3282. doi: 10.1021/jp0138173.
Entropía, glucolisis y cáncer
Durante procesos patolóticos como el cáncer, los patrones de la glucolisis cambian completamente (efecto Warburg). Esto tiene un profundo efecto en el sistema celular. La glucolisis nos permite conectar la idea de autocatálisis y control de la estructura con la entropía. La ruta glucolítica genera entropía y, globalmente, el metabolismo disipa energía y bombea entropía al exterior. La entropía vertida al exterior supera la incorporada al interior (en forma de nutrientes y calor) y la generada por el sistema, resultando en una disminución temporal de la entropía interna. Esto se denomina principio de mínima producción de entropía de Prigogine: el sistema metabolico tiende a minimizar la producción de entropía y maximizar el vertido de entropía al exterior, de modo que el balance favorezca el crecimiento o mantenimiento estacionario de la estructura biológica. Otros científicos piensan mas bien que las células siguen el teorema de máxima producción de entropía. Esto es, la vida evoluciona maximizando el flujo de entropía hacia el exterior. En cualquier caso, la glucolisis es un sistema esencial para la vida, pues es el impulsor, mediante la generación de entropía, del sostenimiento de toda la organización biológica.
En el cáncer se produce un cambio muy profundo en los balances energéticos de las células. La glucolisis cambia durante el desarrollo de tumor. Se ha sugerido que las células tumorales, que, debido a las condiciones ambientales dentro del tumor (menor disponibilidad de oxígeno y competición por la glucosa), favorecen la via de fermentación frente a la respiración, lo que implica una reducción en la producción de entropía. Ello mejora su adaptación a un entorno competitivo mejorando su proliferación y favoreciendo la extensión del tumor. El desarrollo tumoral entonces sería una muestra del funcionamiento del principio de mínima producción de entropía: el tumor evoluciona minimizando su producción de entropía en respuesta a la presión del entorno.
El hidrógeno, el combustible de las células
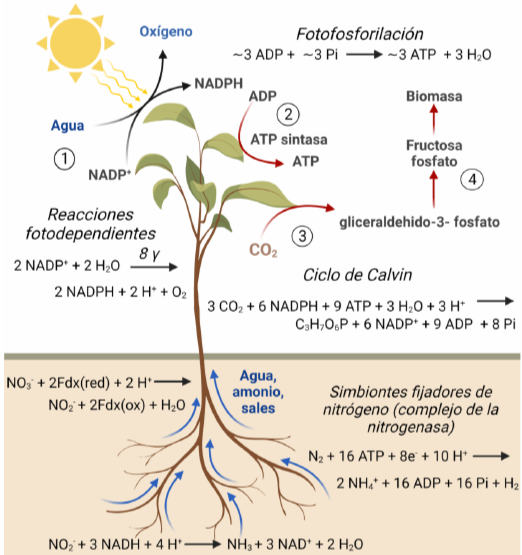
La vida es una estructura que surge durante el proceso de disipación de energía con un bajo flujo de entropía a un alto flujo de entropía. En el proceso es el hidrógeno el que juega el papel energético fundamental. El gran impulso evolutivo en la vida terrestre lo dió la fotosíntesis oxigénica. Los organismos fotosintéticos escinden el agua en oxígeno e hidrógeno, que es una fuente de energía valiosa. La energía del hidrógeno se maneja en forma de cofactores NADH y NADPH.
La fotosíntesis. Los organismos fotosintéticos almacenan energía solar en forma de productos con baja entropía. Ello gracias al hidrógeno, que generan por la ruptura del agua. La biomasa generada alimenta los organismos heterótrofos, que la transforman en hidrógeno de nuevo, que se quema, disipando la energía y aumentando la entropía. Globalmente, se disipa la energía solar con un flujo de entropía neto al exterior que favorece el mantenimiento de la biosfera.
Esta energía se utiliza en la generación de biomasa, que almacena la energía del hidrógeno en forma de productos con baja entropía. La biomasa (particularmente los azúcares) son una buena forma de transportar la energía de unos organismos a otros. Los organismos heterótrofos disiparán la energía, alimentandose de moléculas con baja entropía y generando un flujo de entropía más elevado al exterior. Gracias al metabolismo central, vuelven a recuperar la energía en forma de hidrógeno (NADH y NADPH), que se utiliza para mantener un gradiente electroquímico que se disipa (el hidrógeno se combina con el oxígeno formando agua) y mueve una ‘dinamo’ que produce ATP que, a su vez, controla el mantenimiento de toda la estructura metabólica y molecular. Toda la estructura es temporal y depende del flujo de entropía hacia dentro y fuera del sistema, y de la disipación de la energía. En el momento en el que cambia el balance de entropía, la estructura desaparece: extinción o muerte.
Una vez alguien me preguntaba qué diferencia hay entre un cristal, una estructura de baja entropía, y la vida, una estructura que requiere a su vez baja entropía. La diferencia es que la vida es un proceso. Es una estructura temporal que surge durante la disipación de energía, desde una fuente de baja entropía a un estado de alta entropía.
Como las ondas y formas que surgen durante el proceso de mezclar leche con café, en el que pasamos de un estado de baja entropía (café y leche por separado) a uno de mayor entropía (una mezcla homogénea), así la vida surge temporalmente en el proceso de disipación de un estado de baja entropía (la Tierra primitiva y la energía del Sol) a un estado de alta entropía, en el que esas energías se han ‘esparcido’ en el universo.
Referencias
Bar-Even, A. et al. (2012) ‘Rethinking glycolysis: On the biochemical logic of metabolic pathways’, Nature Chemical Biology. Nature Publishing Group, 8(6), pp. 509–517. doi: 10.1038/nchembio.971.
Baier, G., Müller, M. and Ørsnes, H. (2002) ‘Excitable Spatio-Temporal Chaos in a Model of Glycolysis’, The Journal of Physical Chemistry B, 106(12), pp. 3275–3282. doi: 10.1021/jp0138173.
Kleidon, A. (2009) ‘Nonequilibrium thermodynamics and maximum entropy production in the Earth system’, Naturwissenschaften, 96(6), pp. 653–677. doi: 10.1007/s00114-009-0509-x.
Nielsen, K., SØrensen, P. G. and Hynne, F. (1997) ‘Chaos in glycolysis’, Journal of Theoretical Biology, 186(3), pp. 303–306. doi: 10.1006/jtbi.1996.0366.
Sabater, B. (2022) ‘Entropy Perspectives of Molecular and Evolutionary Biology’, International Journal of Molecular Sciences, 23(8), p. 4098. doi: 10.3390/ijms23084098.
¿Estamos solos en la galaxia?
escrito por C. Menor-Salvan | 3 octubre, 2023
C. Menor-Salván. Agosto 2023
*ver The Conversation para una versión divulgativa corta más actual.
«»A veces creo que hay vida en otros planetas, y a veces creo que no. En cualquiera de los dos casos la conclusión es asombrosa.»»
— Carl Sagan
Esta cuestión lleva resonando en nuestra mente colectiva desde que los humanos comenzaron a observar el Cosmos desde un punto de vista científico, sin invocar dioses ni mitos. Descubrir que no estamos solos en nuestra galaxia o en el Universo sería el mayor y más relevante hallazgo de la Historia humana. Pero, por el momento, solo podemos especular, con más o menos fundamento, acerca de ello.
La pregunta ha vuelto a ponerse de moda a raíz de la reciente declaración de unos ex-militares estadounidenses encabezados por el oficial de la USAF David Grusch, que aseguran que la fuerza aérea de los EEUU oculta en secreto naves extraterrestres y ‘restos biológicos no humanos’ (¿plantas? ¿animales? ¿alienígenas?). Para contextualizar, aquí me entrevistaron sobre ello.
No vamos a hacer una discusión académica sobre la posible vida extraterrestre y la paradoja de Fermi, sino tan sólo delimitar la cuestión (u organizar nuestra ignorancia), hacer algunas reflexiones sencillas al alcance de cualquier lector y dar mi opinión como astrobiólogo de estas ‘nuevas’ (veremos que de nuevas nada) declaraciones. En las referencias se encuentra la bibliografía que he usado y que permitirá al lector interesado profundizar en el tema.
En este artículo, vamos a seguir esta línea de razonamiento:
Acotando la terminología: ‘vida’ vs ‘vida inteligente’
La vida terrestre como modelo, el único que tenemos: cuándo surge la vida y cuándo surge la ‘vida inteligente’.
La vida puede ser común, pero la inteligencia no tiene por qué surgir siempre.
¿cómo buscamos vida extraterrestre?: Sistema Solar y exoplanetas.
¿cómo buscamos vida inteligente? escuchando señales. Una estrategia condenada al fracaso: la galaxia (no ya el universo) es muy vasta. Si están puede que no los escuchemos nunca, ni ellos a nosotros.
La tecnología, ¿podrá resolverlo en el futuro o nos acercamos al final del desarrollo tecnológico?
¿nos han visitado alienígenas?. ¿hay evidencias? ¿es posible una galaxia tipo ‘Star Trek’?
Conclusiones
A qué nos referimos cuando decimos ‘solos en el universo’
Cuando los científicos hablamos de vida extraterrestre no nos estamos refiriendo a ‘vida inteligente’ sino a cualquier forma de vida. Nos interesa descubrir cómo se origina la vida, cómo evoluciona en un planeta, si la vida se originó en otro lugar además de en la Tierra, o si hay lugares que reúnen las condiciones para que surja vida o que se encuentran en la fase de evolución química (lo llamamos ‘habitabilidad’). Todos ellos son parte de los objetivos de la ciencia de la Astrobiología.
Esquema general de la evolución de la vida terrestre y de las aproximaciones para entender su origen.
Usando nuestro planeta como referencia, hay que tener en cuenta que calculamos que la vida se originó aquí hace unos 4200 millones de años. Se calcula que la vida pudo surgir en unos 40 a 120 millones de años desde que comenzó a ser habitable, lo cual es bastante rápido en términos geológicos. Probablemente, una vez dados los primeros pasos, el proceso debió ser muy rápido.
Durante siguientes 2500-3000 millones de años, la vida terrestre estuvo dominada por organismos unicelulares o pluricelulares muy simples (antecesores de las algas actuales). Los animales habitan la Tierra desde hace unos 500 a 600 millones de años. Es decir, los animales han ocupado el planeta durante el último 14% del tiempo desde que existe la vida en éste. Prácticamente somos unos recién llegados.
Durante los primeros 2000 millones de años de vida en la Tierra la situación era mas o menos así: cianobacterias. Si encontrásemos vida extraterrestre, lo más probable es que se parezca más a esto que a ET o Predator.
Dado que el concepto de inteligencia, como el de vida, es algo difuso, definiremos como ‘vida inteligente’ aquella con la capacidad de observar el cosmos.
¿cuándo surgió en la Tierra una forma de vida que comenzó a poner sus ojos en el firmamento y a andar el camino hacia su exploración?.
Si ponemos la Edad de Bronce como el inicio de las primeras observaciones astronómicas sistemáticas, digamos que tenemos vida inteligente en la Tierra capaz de observar el Cosmosdurante el último 0.00000012% del tiempo de existencia de vida. Los científicos también hablan de vida inteligente con tecnología de comunicación capaz de mandar señales fuera del planeta. Los humanos somos capaces de ello desde la invención de la radio, es decir, desde hace poco más de un siglo.
Cálculos recientes, basados en el análisis bayesiano de los datos de la vida que conocemos, sugieren que hay aproximadamente un 60% de probabilidades de que nunca surja vida inteligente en un planeta con vida. Es decir, la evolución de la vida no necesariamente va a conducir a que emerja vida inteligente. Si la vida es una consecuencia de la evolución del cosmos, no tenemos razones para pensar que la inteligencia también lo sea.
Con estos datos, es fácil asumir que, si encontramos evidencias de vida extraterrestre, seguramente serán de vida simple (unicelular o pluricelular) o, en general, vida no inteligente. Como es lógico, no necesariamente tiene que ser así, pues no conocemos cuánto tiempo puede tardar en emerger vida inteligente en un planeta; en la Tierra puede que haya tardado mucho, dada la cantidad de eventos de extinción que han ocurrido.
O bien, al contrario, puede que el impulso evolutivo que dan las crisis biológicas haya hecho que surja más pronto. No hay forma de saberlo aún. Pero, con lo que conocemos, no es descabellado asumir que, probablemente, la vida sea frecuente en la galaxia, pero que la vida inteligente sea bastante poco común o incluso única. De hecho, otros cálculos recientes sugieren que, en nuestra galaxia, podría haber entre 1 y 10 planetas con vida inteligente capaz de explorar el cosmos, incluyendo el nuestro.
Los últimos 500 millones de años de vida en la Tierra son los más animados desde el punto de vista biológico. La barra está a escala; obsérvese cuando surgen los humanos.
En este sentido, hay que aclarar que la ecuación de Drake (formulada a partir de una famosa pregunta de Fermi a sus estudiantes: ¿cuantos pianos hay en Chicago?) no pretende ser una estimación precisa, sino una aproximación mental y una delimitación de nuestra ignorancia. El propio Drake solía responder que el número de civilizaciones está «entre una y mil millones».
Entonces, ¿no tenemos evidencias de vida extraterrestre?
No, no tenemos ninguna evidencia. Buscamos evidencias de vida, es decir, biofirmas, de varias formas:
Huellas moleculares, geoquímicas o físicas. Estas pueden ser biomarcadores orgánicos o inorgánicos, es decir, compuestos químicos que estén ligados a la vida. Pueden ser huellas geoquímicas o geológicas, tales como determinados minerales o huellas isotópicas.
También pueden ser señales espectroscópicas o biofirmas basadas en fenómenos químico-físicos, tales como desequilibrios químicos, estructuras termodinámicas lejos del equilibrio.
O, simplemente, pueden ser indicios de habitabilidad, tales como presencia de agua líquida o temperaturas en el rango de estabilidad de moléculas orgánicas
Buscamos todas estas biofirmas, bien de vida o bien de evolución química, e indicios de habitabilidad en otros lugares de nuestro Sistema Solar que pudieron ser habitables (Marte) y en los exoplanetas, planetas descubiertos en otros sistemas planetarios.
La búsqueda de biofirmas en exoplanetas aún es un campo en el que hay mucho que desarrollar y se basará fundamentalmente en la búsqueda de señales espectroscópicas que indiquen habitabilidad o vida y que nos permitan conocer la composición atmosférica en cierto detalle.
Los astrónomos están investigando exhaustivamente las estrellas que nos rodean, nuestro vecindario cósmico, buscando nuevos planetas. El catálogo de exoplanetas de la ESA y el catálogo de la NASA indican que, mientras escribo estas líneas, se han identificado nada menos que 5483 planetas en 4220 sistemas planetarios, todos en nuestra vecindad galáctica. Esta lista incluye planetas desde tamaños mucho mayores que Júpiter a tamaños similares e incluso menores a la Tierra. Algunos eran prometedores candidatos a ser planeta habitable.
Sin embargo, al profundizar en su observación, todavía no hemos encontrado ningún planeta que sea un serio candidato a reunir condiciones de habitabilidad o vida. En los planetas cuya temperatura han podido estimar los científicos, todos salvo uno son demasiado calientes para sostener vida orgánica. Y el único con temperatura moderada, es un gigante gaseoso mayor que Júpiter.
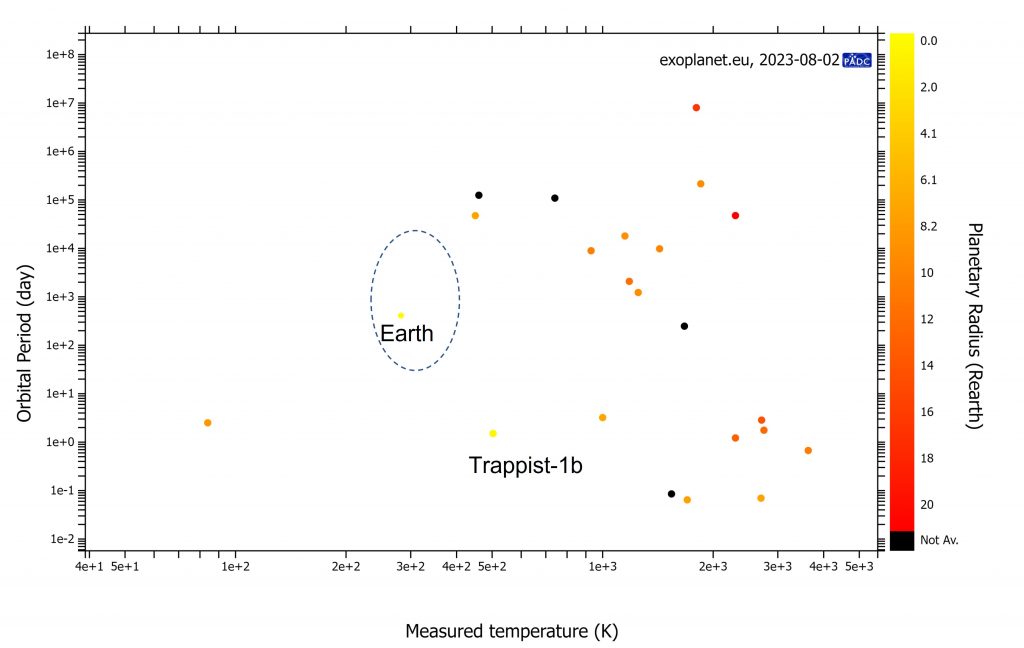
Scatter plot de los exoplanetas cuya temperatura ha podido determinarse hasta la fecha de hoy. La temperatura mas baja, 84K, se corresponde al planeta Qatar-3b, situado en el sistema Qatar-3, una estrella similar a nuestro Sol situada a más de 2300 años luz. El planeta es un enorme ‘Júpiter caliente’ y la estrella es demasiado joven (0.3 Ga), así que la vida está prácticamente descartada en ese sistema. Hemos ubicado el lugar donde estaría la Tierra en este gráfico y la región donde esperamos planetas con posible vida (circulo). Trappist-1b y 1c, a 40 años luz de la Tierra, fueron muy prometedores cuando se descubrieron; la medida de su temperatura da un valor medio de 503K, demasiado cálido para sostener vida orgánica, además de estar en bloqueo de marea y la proximidad a su estrella, lo que sugiere alta irradiación. Trappist-1b tiene una atmósfera densa rica en agua, lo que sugiere que en el pasado pudo tener océanos y ahora es demasiado caliente (efecto invernadero desbocado).
La mayor parte de los exoplanetas identificados son demasiado calientes, sometidos a intensa radiación, están en acoplamiento de marea o, en general, tienen características que descartan su habitabilidad; pero, la mayoría de los sistemas planetarios tienen edades comparables al nuestro y muchos tienen múltiples planetas. La presencia de planetas habitables (y no digamos habitados) no parece demasiado común, al menos de momento y en nuestro vecindario cósmico, pero no perdemos la esperanza y, posiblemente, con la mejora de la tecnología de observación tengamos sorpresas pronto.
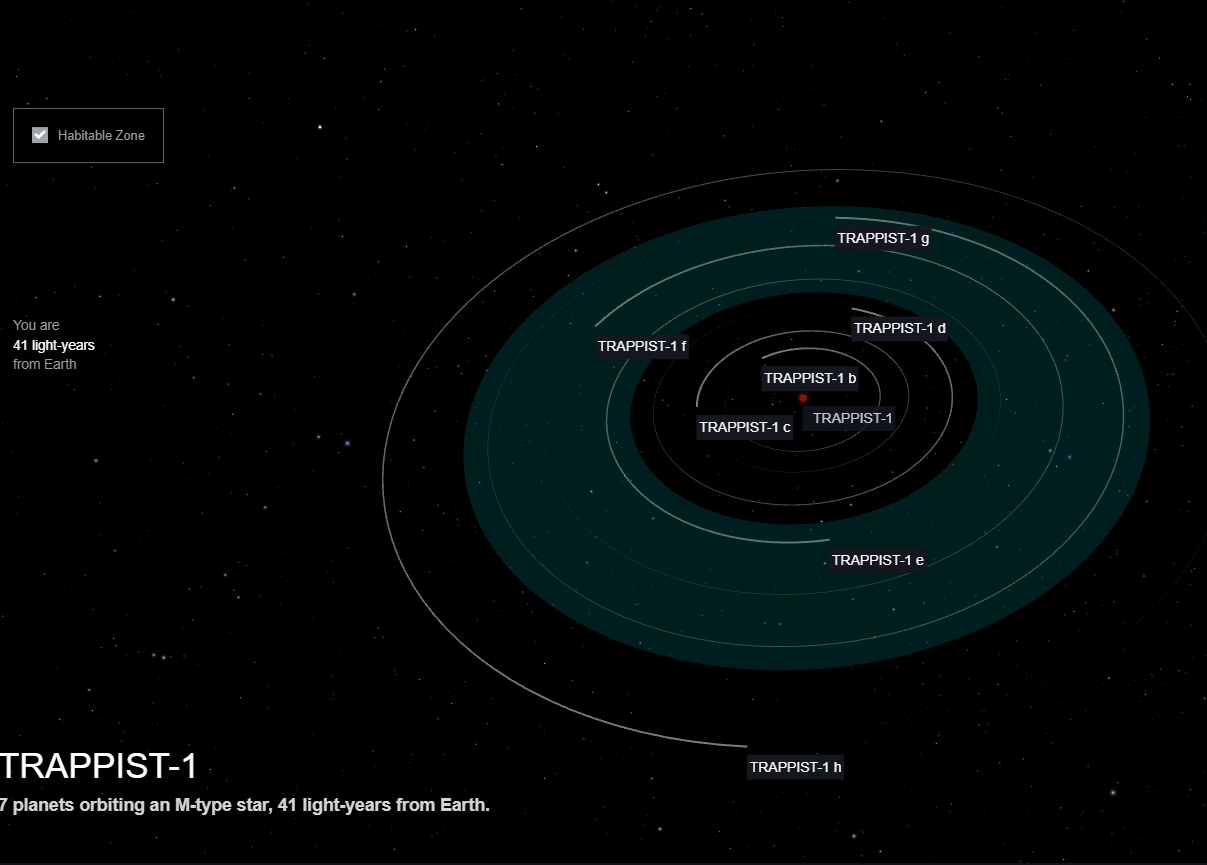
El sistema Trappist-1, a tan sólo 41 años luz de nosotros, contiene 7 planetas, varios de ellos rocosos y de tamaños comparables a la Tierra. Su descubrimiento causó gran agitación: ¿se encontró un sistema planetario con posible vida?. Varios de sus planetas están en la región de habitabilidad. Sin embargo, su estudio detallado sugiere que ninguno de los planetas es habitable, aunque todavía no hay nada concluyente. Se ha sugerido que la estrella tiene una edad avanzada y que varios de sus planetas, como el 1b y 1c, pudieron ser habitables en el pasado. Actualmente, 1f podría ser un mundo helado similar a Europa o Encelado. No obstante, no hay nada cerrado y futuras observaciones pueden darnos sorpresas.
Pero, por el momento, no hay evidencias de vida extraterrestre y ni siquiera podemos confirmar que ninguno de los planetas observados sea habitable en el sentido astrobiológico. Ya podemos ir haciendo números con lo que sabemos: se han observado unos 1800 planetas rocosos (tipo terrestre o ‘supertierras); aunque hayan surgido sospechosos, no podemos confirmar que haya vida o condiciones de habitabilidad en ninguno de ellos.
Ello implica que, de momento, hay una probabilidad de un 99.94% de que un planeta rocoso tipo terrestre recién encontrado no tenga vida. Encontrar un planeta habitable es algo parecido a que te toque la lotería. La diferencia es que nuestra galaxia juega muchos números.
Pero, no sólo debemos tener en cuenta los planetas. Se han observado 849 planetas con masa mayor que la de júpiter. Estos planetas pueden tener lunas con posibilidades de habitabilidad, como, en nuestro sistema solar, es el caso de Europa o Encelado. Aún no hay capacidad tecnológica para observar las exolunas, aunque ya hay algún candidato a exoplaneta con sistema de lunas. Ello multiplica las posibilidades de que exista vida en nuestro entorno planetario.
Impresión de cómo podría ser la exoluna Kepler-1625b-i, que orbita el planeta Kepler-1625b, un ‘superjúpiter’ de aproximadamente el tamaño de nuestro júpiter pero mucho más denso. Confirmar la existencia de exolunas es muy difícil por el momento.
Por ello, debemos seguir investigando, pues queda mucho espacio por observar y aún tienen que mejorar mucho las observaciones. Puede que en un futuro próximo alguno de los observados, o uno nuevo, pueda confirmarse adecuado para el origen o presencia de la vida.
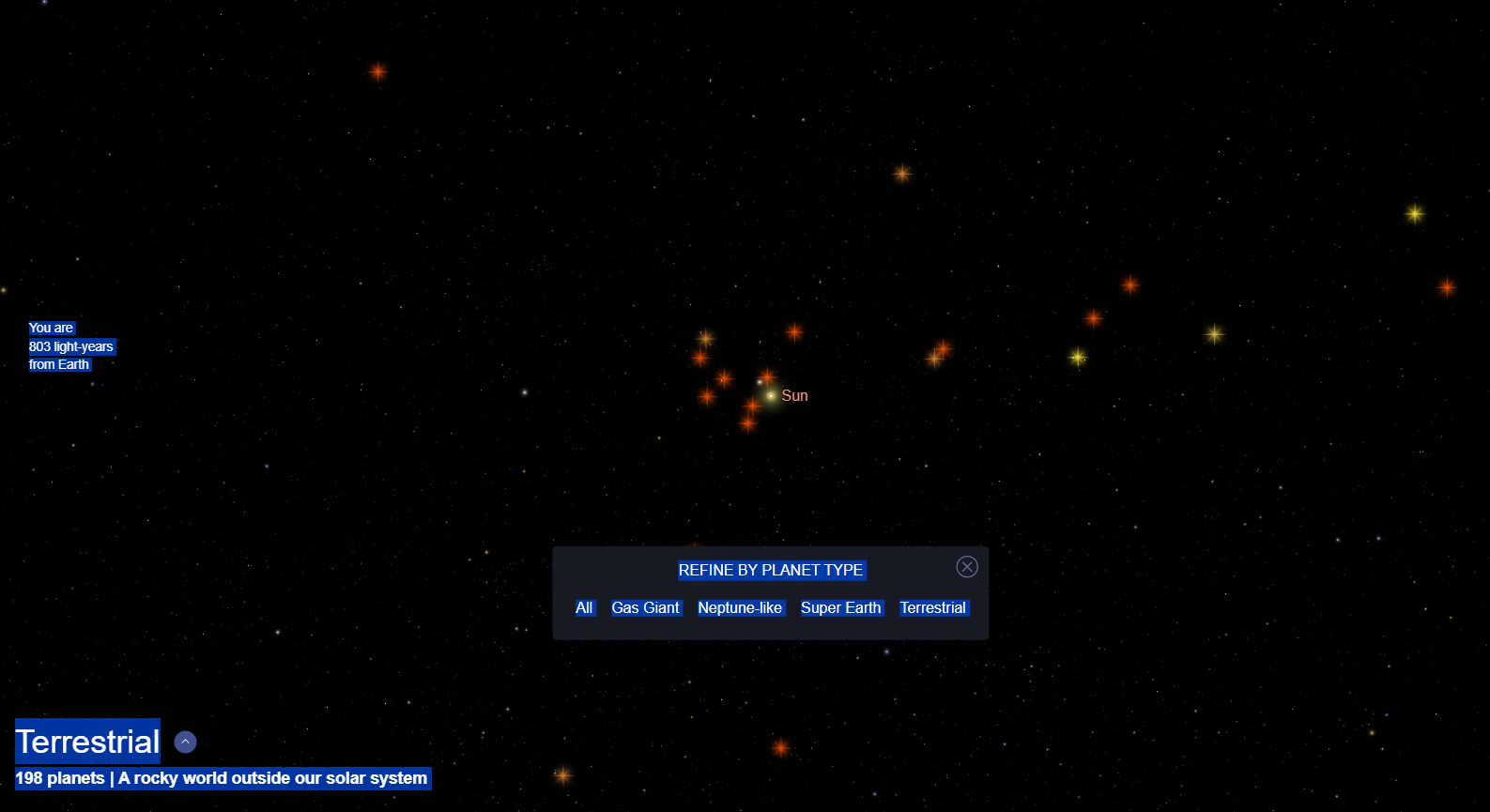
Mapa en 3D de los sistemas con planetas de tipo terrestre (rocosos y tamaños similares a la Tierra) descubiertos por ahora en nuestro vecindario galáctico, en el precioso catálogo de exoplanetas de la NASA. De todos los planetas identificados, 198 son de tipo terrestre pero ninguno de ellos parece ser habitable. De momento no hay vulcanos ni klingons ahí fuera.
¿la vida podría ser frecuente en la galaxia o no?
Pensamos que la formación de vida puede ser relativamente común por varias razones: la universalidad de la química prebiótica, que da lugar a la formación de un set similar de compuestos orgánicos en muchas condiciones y que hemos observado en asteroides, por ejemplo. Es decir, la formación de los precursores orgánicos simples de la vida es un proceso que se integra dentro de la geoquímica de modo normal y, seguramente, es muy corriente en la galaxia.
Por tanto, el primer paso hacia la vida orgánica debe ser común. Tal vez el siguiente paso, el proceso de evolución química, sea menos común. Pero no tenemos motivos para pensar que sea una rareza, ya que es un proceso que se integra en la geoquímica planetaria.
Por otro lado, la evolución de la vida desde un punto de vista termodinámico parece una consecuencia de la propia evolución de los sistemas planetarios. La vida es un eficiente sistema de generación y disipación de entropía. La entropía del universo aumenta, la energía de las estrellas se disipa y, en el proceso, surgen estructuras que podrían maximizar la producción de entropía. La vida surgiría, entonces, como una estructura dinámica en el conjunto de fluctuaciones generadas por los procesos de disipación de energía y aumento de entropía del universo, es decir, una consecuencia del segundo principio de la termodinámica.
Pero, si la evolución y vida parece un imperativo de la propia termodinámica del universo, entonces, ¿por qué todavía no hemos visto ningún planeta habitable o con pistas de vida?.
Por varias razones: aunque la vida sea probable, hemos observado sólo una muestra mínima de planetas. Se calcula que solo en nuestra galaxia hay unos 100.000 millones de planetas. En nuestro vecindario, hasta 50 años luz de distancia, hay unos 1500 planetas; se han observado aproximadamente el 10% de ellos. Es muy probable que la vida sea frecuente en números absolutos, aunque la probabilidad de encontrar un planeta con vida o habitable sea baja; tan sólo hemos observado el 0.000005% de los planetas de nuestra galaxia.
No puede decirse ni que tengamos mala suerte buscando, pues ni siquiera conocemos al 90% de los vecinos de nuestra propia manzana.
Si nosotros no los vemos, ¿pueden vernos ellos?
Si hay un planeta con vida inteligente en un radio de unos 3000 años luz, con una tecnología de observación similar o superior a la nuestra, cabría la posibilidad de que hubieran encontrado nuestro Sistema Solar y catalogado nuestro planeta como habitable o con vida, en base a las huellas espectroscópicas, distancia del Sol, masa, movimiento y otras características.
Esto podría ocurrir ahora (lo cual es extremadamente improbable), o haber ocurrido en algún momento durante los últimos cientos o miles de millones de años. O podría ocurrir en un futuro. No hay nada que descarte que nuestro planeta ha sido ya observado.
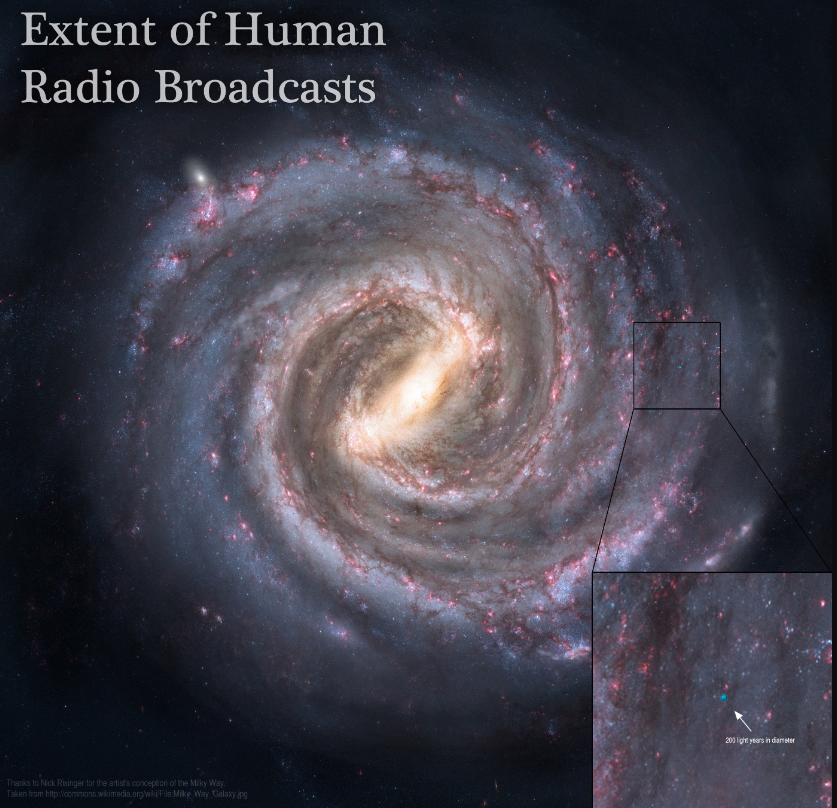
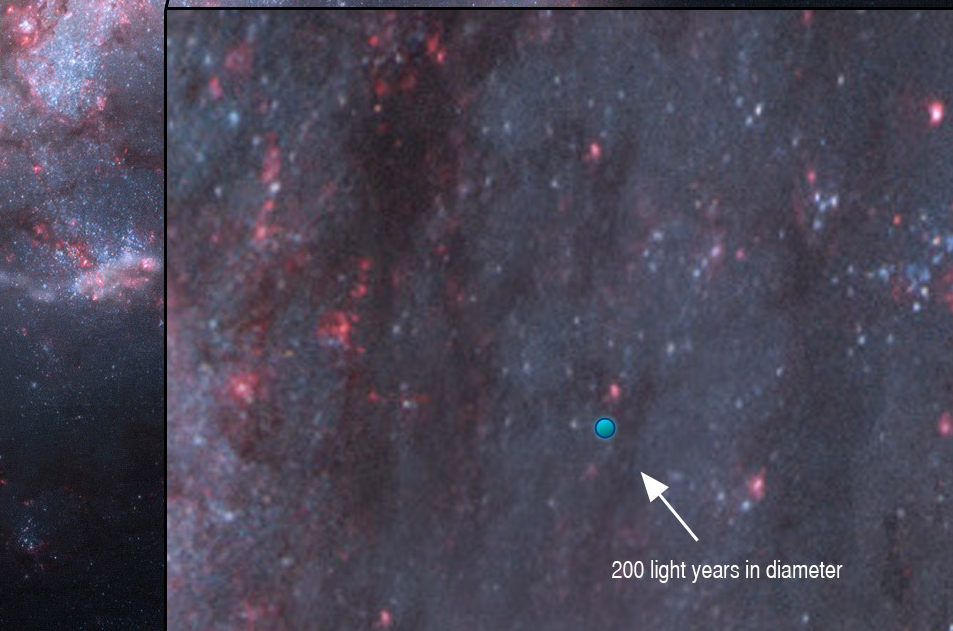
En cuanto a detectar nuestras emisiones de radio y determinar que provienen de seres inteligentes, es mucho menos probable. Teniendo en cuenta que llevamos emitiendo señales al espacio unos pocos años, estas serían detectables en una pequeña burbuja de unos 200 años luz de diámetro. A través del proyecto SETI, los humanos llevan varios años buscando débiles señales que sugieran vida inteligente. Nunca se recibió ninguna. Una vez hubo un susto, la famosa señal wow, pero no hubo suerte.
No obstante, llevamos poco tiempo escuchando. Sería como encender la radio durante un segundo, justo cuando no se oye a nadie hablando, y pensar que no hay nadie ahí. O, como nos ocurre a nosotros, la extensión de la burbuja de emisiones de una civilización extraterrestre quizá quede demasiado lejos. Es como tratar de comunicarnos con alguien usando un walkie talkie o una pequeña emisora de poco alcance. Si no hay nadie con un receptor, nadie va a oir nuestra emisión ni responder. Pero ello no implica que no haya nadie en los alrededores.
Extensión de nuestras emisiones de señales: el pequeño punto azul, insignificante en la galaxia. Imagen de Adam Grossman y Nick Risinger, que puede verse en alta resolución, aqui en The Planetary Society.Detalle de la burbuja azul de nuestras emisiones de radio. Detectar emisiones de seres extraterrestres implicaría que su burbuja nos ha alcanzado. Esto es muy poco probable, aunque hayan estado emitiendo.
Además hay que tener en cuenta que las señales electromagnéticas (radio, luz,…) disminuyen su intensidad de modo proporcional a 1/d2, con lo que necesitan instrumentación sensible. La probabilidad de que haya una civilización o inteligencia extraterrestre buscando señales extraterrestres (para ellos) con instrumentación sensible, en un área conteniendo unos 6000 planetas, es ínfima, máxime cuando los cálculos más optimistas sugieren que, en este momento, la galaxia puede contener unos 10 planetas con vida inteligente capaz de explorar el cosmos.
Así que, quizá estén ahí fuera y, ni nos oigan, ni les oigamos jamás. O quizá surja en el futuro una civilización que detecte una débil señal procedente de nuestro planeta, donde una civilización desapareció miles de años antes.
Quizá sea una cuestión de limitación tecnológica: el gran filtro
Aquí la imaginación es el límite: podemos imaginar que una inteligencia extraterrestre ha encontrado formas de escuchar señales lejanas o viajar a otros puntos de la galaxia. Pero lo que podamos imaginar no nos sirve si no lo podemos sustanciar en una aproximación científica.
Podemos afirmar que quién sabe qué tecnologías desarrollaremos. Este pensamiento es muy común: «hace 100 años apenas éramos capaces de volar, y ahora salimos al espacio. ¡Quién sabe qué haremos en 100 años más!»
Esta idea presupone una extrapolación lineal en el desarrollo de la tecnología. La mente humana tiende a pensar de modo lineal: si en 100 años hemos conseguido tal cosa, en otros 100 extrapolamos esto linealmente y llegamos al mundo de Star Trek.
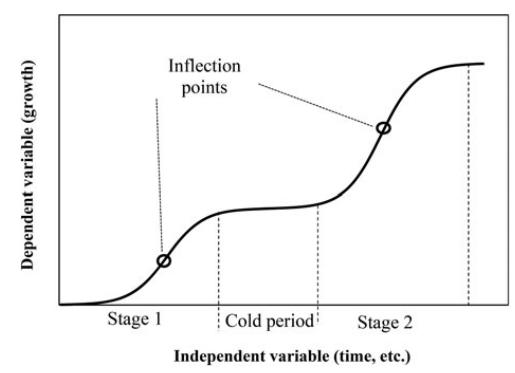
El problema es que la Naturaleza no suele seguir funciones lineales. El desarrollo tecnológico humano, como tantos procesos de desarrollo o crecimiento, sigue una función sigmoidal. Voy a usar como analogía el crecimiento de la colza:
El conocimiento y la tecnología siguen un patrón similar. Tras una fase inicial lenta, se llega a una fase de aceleración exponencial, seguida de una fase de meseta, en la que el rápido incremento en el conocimiento y la tecnología se frena. Aquí pueden ocurrir dos cosas: que termine declinando la civilización y se pierda el conocimiento y la tecnología, o que haya un ‘salto evolutivo’, una nueva tecnología o cambios de paradigmas que permitan un nuevo periodo de crecimiento. ¿Dónde estamos nosotros ahora?. ¿llegando a un punto de inflexión? ¿entrando en un periodo frío? No se si alguien lo habrá estudiado, pero no he encontrado referencias. Así que, teniendo en cuenta las funciones sigmoidales, dado que hemos tenido un rápido crecimiento tecnológico y en conocimientos, en algún momento se llegará a una meseta, si no estamos llegando ya.
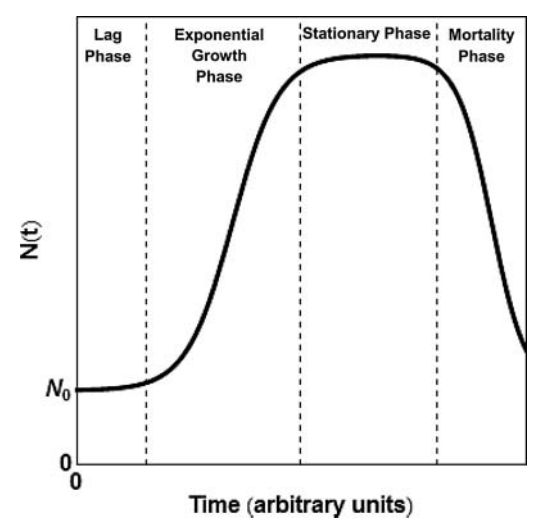
Teniendo en cuenta que todos los fenómenos de crecimiento y poblacionales siguen una función sigmoidal (ya sea el crecimiento de una colonia de bacterias, de un bosque, de la capacidad de computación, de una población, de una pandemia, de la economía….), la idea del ‘gran filtro de Hanson‘ es consistente con un modelo sigmoidal simple: podemos sugerir que toda civilización inteligente llega, tras un periodo de rápido crecimiento tecnológico, a una meseta, seguida de decaimiento y, eventualmente, desaparición de esa civilización. Dado que es una ley natural de los procesos de desarrollo poblacionales, con seguridad una civilización extraterrestre basada en poblaciones de individuos va a seguir ecuaciones sigmoidales.
Todo crecimiento en un sistema cerrado (ya sean bacterias en un cultivo o una civilización) siguen una curva sigmoidal que llega a una meseta y decaimiento (extinción). Este modelo es consistente con la idea del ‘gran filtro’
La idea del gran filtro implicaría que, en un planeta, como sistema semiabierto (como es por ejemplo un cultivo bacteriano), toda civilización llegaría a la fase de extinción antes de que tengamos la posibilidad de encontrarnos. La única forma de evitarla (como en un cultivo bacteriano) es contactar con el exterior y ‘pasar’ parte de la población a un nuevo sistema (pasando bacterias a un nuevo cultivo). Entonces, si no existe la posibilidad física de sortear la distancia o de encontrar ninguna otra de esas civilizaciones o inteligencias potenciales, no nos queda otra que extinguirnos en silencio. Un mundo como el de Star Trek resolvería el problema del gran filtro; implicaría que hay un salto tecnológico, de conocimiento o evolutivo que no podemos imaginar y que nos llevaría a un nuevo periodo de crecimiento exponencial, hasta el siguiente punto de inflexión.
En Star Trek: First Contact, el hallazgo del imaginario ‘motor de curvatura’ y la llegada de los vulcanos salva a los humanos de caer en el gran filtro.
Entonces, ¿nos ha visitado una inteligencia o inteligencias extraterrestres? ¿qué hay de los recientes testimonios sobre alienígenas en el Congreso de los EEUU?
Las noticias y testimonios sobre el fenómeno OVNI y visitas extraterrestres han existido desde el siglo XIX. Periódicamente, el mito de los misteriosos hangares y centros de investigación estadounidenses con naves extraterrestres y restos de alienígenas se reaviva. Surgen oscuros testimonios, vagas explicaciones, imágenes borrosas que sirven, sobre todo, para hacer películas de ciencia ficción sobre el tema.
Pero, no hay ni una sola prueba.
Los científicos no trabajamos con anécdotas ni testimonios. Trabajamos con evidencias. Usamos la ‘Navaja de Hitchens’:
una afirmación sin evidencias se puede descartar sin evidencias»
Los científicos (los periodistas, en teoría, también. Christopher Hitchens era periodista) simplemente descartamos los testimonios que no se sustentan en pruebas, datos y evidencias. Y en afirmaciones como esta se requieren datos sólidos. Como dijo Carl Sagan, afirmaciones extraordinarias requieren pruebas extraordinarias.
David Grusch, uno de los declarantes en el Congreso sobre los UAP y la supuesta tenencia de tecnología y restos biológicos extraterrestres. Una declaración ridícula, hueca y, desde el punto de vista científico, totalmente descartable. Personalmente, me pareció todo una broma.
Estas recientes declaraciones en el congreso de EEUU han sido un compendio de los clásicos testimonios sobre fenómenos OVNI que hemos visto en otras ocasiones: nada concreto, ninguna prueba, evasivas e incluso afirmaciones, digamos, interesantes, como que el declarante sugiere que los extraterrestres estaban muy interesados en la tecnología nuclear. ¿una inteligencia con tecnología tan avanzada como para cruzar cientos o miles de años luz, interesada en una tecnología seguramente obsoleta? ¿tal vez quiere decir que la energía nuclear es muy importante y en su planeta se quedaron sin uranio?.
El testimonio era incluso ridículo cuando le preguntan al declarante si había visto personalmente esos restos de naves alienígenas y restos biológicos ‘no humanos’ y dice que no personalmente, que se lo han contado. No sólo declaran sin aportar pruebas, sino en base a rumores.
El ‘black mailbox’, en Nevada. En la ruta de los entusiastas del ‘fenómeno OVNI’, próxima al área 51. Aquí esperan ver o recibir alguna comunicación de los extraterrestres.
Estas declaraciones ni siquiera pueden considerarse ciencia ficción, sino meras elucubraciones y rumores. Además, estas cosas no tienen en cuenta algo elemental sobre la naturaleza humana: no sabemos guardar secretos. ¿un centro de investigación con naves extraterrestres capturadas, lo que sería el hallazgo más importante de la Historia de la Humanidad, por el que habrían pasado miles de trabajadores, científicos y militares, oculto y secreto desde los años 1930?.
Este nuevo episodio de declaraciones sobre supuestas tecnologías extraterrestres me recuerda al caso de Bob Lazar. Este curiosísimo personaje afirmó haber trabajado en el ‘área 51’ y en un lugar secreto llamado S-4, donde se realiza ingeniería reversa de naves extraterrestres, y realizó declaraciones muy similares a las que se han escuchado estos días en el Congreso de los EEUU. Afirmó que unos seres extraterrestres provenientes de Zeta-Reticuli, un sistema estelar situado a unos 39 años luz, utilizaban en sus naves el ‘elemento 115’, de propiedades muy inusuales. Bob Lazar llegó a ser muy popular en el colectivo de los conspiranoicos y fans del fenómeno OVNI. Sin embargo, el descubrimiento de que zeta-reticuli carece de planetas en su sistema y la síntesis del elemento 115 en 2003 demostraron que las afirmaciones de Bob Lazar no eran más que o delirios, o bien un plan para ganar notoriedad y hacer su negocio en torno al mundo OVNI.
Bob Lazar con uno de sus esquemas de naves extraterrestres. ¿embaucador, perturbado o visionario?
Bob Lazar es un personaje curioso no obstante. Siempre me preguntaré de dónde obtuvo muestras de plutonio (cantidad ínfima, obviamente), de las cuales estuve a punto de adquirir una para mi colección cuando fue detenido por el FBI.
Los científicos debemos mantener la mente abierta. Debemos investigar. Pero investigamos siguiendo una serie de metodologías que evitan que caigamos en las trampas cognitivas comunes o terminemos cayendo en las redes de embaucadores como Lazar o David Grusch. Richard Feynman decía que la persona más fácil de engañar era uno mismo. La Ciencia evita que caigamos en nuestras propias trampas mentales. Hasta que no haya pruebas, este tipo de declaraciones no son más que ruido.
¿Debemos investigar los fenómenos anómalos?
Por supuesto. No hay que confundir los reportes de UAP con las declaraciones sobre objetos extraterrestres en oscuras instalaciones militares. Los reportes de fenómenos anómalos están recogidos por instrumentos y radares de aeronaves y son datos objetivos. Otra cosa es si podemos averiguar la causa o no.
¿por qué somos científicos? movidos por la curiosidad, la necesidad de explorar lo que no conocemos, de responder a cuestiones fundamentales que ya nos hacíamos de niños: ¿por qué la hierba es verde, por qué podemos respirar?. Cuando voy a museos o ferias de minerales, siempre hay niños llenos de curiosidad. Pocos niños hay a quienes no les encanten los minerales. Suelen hacer observaciones muy interesantes: ven los aspectos en común que tienen minerales con composición similar, o reconocen las formas cristalográficas. Esa misma curiosidad es la del científico. Lamentablemente, la sociedad tiende a desanimar a esos niños, a destruir su curiosidad. Un científico es alguien que mantuvo esa faceta infantil. De hecho, la curiosidad es el principal, pero el menos valorado activo que posee un científico. Así que, ¿cómo nos resistiríamos a investigar y explorar cualquier fenómeno extraño o anómalo?. No seríamos científicos entonces.
Pero no investigamos de cualquier forma, ni nos dejamos arrastrar por vagos testimonios. Ni afirmamos, ni descartamos nada, sino que planificamos cómo abordar una cuestión o problema. Adoptamos la postura escéptica en el sentido clásico del término, la epojé de Sexto Empírico: suspensión de juicio u opinión durante la observación y experimentación. Por lo que hemos visto anteriormente, es muy poco probable que coincidamos en el tiempo y en la vecindad galáctica con otra forma de vida inteligente. Es poco probable que ésta haya alcanzado un nivel tecnológico suficiente y es poco probable que las propias leyes de la Física permitan tales viajes. Pero no lo descartamos. Simplemente, seguimos trabajando y seguimos las evidencias.
En 2022 la NASA creó una comisión para el estudio de los UAP o ‘fenómenos anómalos no identificados’ (unidentified anomalous phenomena). Este es el nombre correcto, no ‘fenómenos aéreos no identificados’, como indicaron algunos medios, denominación incorrecta que se abandonó.
Estos fenómenos UAP engloban supuestos avistamientos OVNIS (hay cientos de relatos de pilotos en torno a ellos) así como cualquier fenómeno, atmosférico o espacial, no identificado.
La comisión independiente para el estudio de los UAP
Los astrobiólogos debemos estudiar todo fenómeno no explicado o anómalo. No se trata de dar oídos a testimonios, mitos o conspiraciones, sino investigar seriamente qué hay. La mayor parte de los UAP tienen explicaciones sencillas. En muchos casos se han cotejado con tráfico aéreo comercial y se corresponden con detecciones de aviones lejanos.
Aun así, hay fenómenos que, por falta de evidencias o datos, no pueden no ya explicarse, sino ni siquiera estudiarse. Ello no implica que existan inteligencias extraterrestres. Que algo no tenga explicación no implica que sea válida cualquier explicación por extravagante que sea. Por ejemplo, que no pueda explicar el Origen de la Vida no implica que el Origen de la Vida sea obra de Dios. Del mismo modo, que no pueda explicar un borroso objeto que un piloto vio una vez en el aire, no implica que fuera una nave extraterrestre.
La falta de datos, simplemente, implica que, o bien hay que seguir estudiando y obteniendo datos, o, si no podemos tener más datos, quedando la cuestión o hipótesis fuera de toda posibilidad de verificación y de falsación (otra de las herramientas del trabajo científico), la tenemos que dejar archivada. Pero, en la práctica, como decía Sherlock Holmes «en general, cuanto más extravagante es una cosa, menos misteriosa suele resultar»
¿Es posible el mundo de Star Trek?
Como científico debo razonar y usar datos y evidencias. Pero no dejamos de soñar. Desde pequeño, cuando pasaba noches enteras mirando el firmamento, primero con los ojos desnudos, luego con mi pequeño telescopio, siempre soñaba con que alguien más habría en algún planeta ahí arriba. Deseaba que vinieran. Eso implicaría que es posible viajar por el espacio y a otros sistemas planetarios. Si realmente hubiera extraterrestres, ello indicaría que nuestro pequeño mundo podría hacerse mucho más grande. Es una gran esperanza. Pero ello queda fuera del ámbito de la Ciencia.
Que triste sería que no hubiera nadie más en nuestra galaxia. Alguien dijo una vez que somos el Universo contemplándose a sí mismo. Resulta difícil aceptar la idea de que, en 100.000 millones de planetas en nuestra galaxia, no haya otras inteligencias planteándose las mismas preguntas. Pero, más difícil aún es aceptar que, habiendo otras inteligencias, estamos aislados en esta isla planetaria. Así que, como todos los que crecimos con Star Trek (yo lo hice con TNG), soñábamos en cada capítulo que podríamos explorar la galaxia en busca de nuevos mundos y nuevas civilizaciones y que igual, por fin, esa noche se iban a presentar visitantes extraterrestres a resolver nuestras dudas.
Siempre podemos soñar que es posible explorar otros mundos como en Star Trek
Pero, ¿es posible el mundo de Star Trek?. Ello tiene varios problemas: el problema de la probabilidad de vida inteligente, el problema de la tecnología y el problema de la coincidencia de civilizaciones en la misma región del universo que hemos revisado anteriormente.
Pero tiene otro problema: un mundo como el de Star Trek presupone que la evolución no es un fenómeno de caos determinista, sino que es teleológica y que termina en seres humanoides que, incluso, pueden cruzarse formando híbridos. Naturalmente, esta trampa teleológica no tiene sentido desde el punto de vista biológico. En cierto modo, Star Trek implica una profunda aceptación del principio del Diseño Inteligente de origen divino, que culmina en seres a imagen y semejanza de Dios (aunque en Star Trek, en lugar de Dios, se apela a unos seres que sembraron, mediante panspermia de su ADN, los mundos habitables). Dentro de la lógica de Star Trek, es casi un imperativo llegar al conocimiento del viaje interestelar. Pero, desde el punto de vista científico, no hay ninguna razón que sugiera que tales viajes serían tecnológicamente posibles.
Si hay vida en otros planetas, es muy posible que sea similar a nivel molecular, teniendo en cuenta que partimos del mismo espacio químico y las reglas de la química imponen cierto determinismo molecular. Pero, igual que en nuestro planeta, donde el mismo patrón molecular y células similares han dado lugar a innumerables morfologías y posibilidades, no podemos ni imaginar cómo sería una inteligencia extraterrestre. No siquiera sabemos si podríamos identificarla con las herramientas de que disponemos actualmente.
La fauna de Ediacara, que desapareció hace unos 540 millones de años, estaba formada por organismos extraños, como este Hallucigenia. Si la misma biología molecular daba lugar a organismos como éstos, ¿cómo serán los organismos extraterrestres? ¿serán molecularmente similares, aunque morfológicamente inimaginables?
Personalmente, desearía que las declaraciones de David Grusch en el congreso fueran reales. Ojalá haya naves alienígenas, lo que implicaría que es posible encontrarnos y explorar los exoplanetas. Pero no tenemos ni una sola prueba o evidencia. Aquí, tras todo lo dicho, sólo veo dos posibilidades: o nos espera un futuro apasionante o una soledad desoladora.
Conclusiones
La vida es probablemente bastante común en nuestra galaxia, pero la vida inteligente es, probablemente, muy rara o incluso única (nosotros) en este momento.
Por ‘común’ queremos decir en números absolutos. Con lo que conocemos a través de la observación de exoplanetas, la probabilidad de hallar un planeta habitable o con vida es muy baja. Es una situación comparable a la lotería: En todo el país va a tocar el gordo de la lotería seguro en algún sitio, pero la probabilidad de que te toque a tí, en tu barrio o en tu ciudad es baja. Con lo que sabemos por el momento, hay un 99.94% de probabilidad de que un planeta terrestre no tenga vida. Lo que pasa es que la galaxia tiene miles de millones de planetas.
Es extremadamente improbable que exista vida inteligente en nuestro vecindario cósmico o nuestra galaxia, y aún más improbable que dispongan de capacidad para realizar viajes interestelares
Las declaraciones sobre OVNIS y UPA no están respaldadas con ninguna evidencia, solo son rumores. La mayoría de los avistamientos documentados por pilotos y registrados por radares, o bien han sido explicados, o bien no hay datos suficientes como para poder estudiarlos. Los científicos no trabajan con testimonios, sin datos suficientes o incompletos. Esto no quiere decir que no se estudie. Hay que revisar objetivamente todos los datos y documentos.
Aun así, todos nuestros cálculos se basan en la vida terrestre como modelo. Esto podría cambiar, por lo que seguimos estudiando e investigando y nos mantenemos alertas.
Referencias
Ćirković, M. M. (2013) ‘Who are the SETI sceptics?’, Acta Astronautica, 89, pp. 38–45. doi: 10.1016/j.actaastro.2013.03.012.
Dick, S. J. (2012) ‘Critical Issues in the History, Philosophy, and Sociology of Astrobiology’, Astrobiology, 12(10), pp. 906–927. doi: 10.1089/ast.2011.0786.
Haqq-Misra, J., Kopparapu, R. K. & Schwieterman, E. (2020) ‘Observational Constraints on the Great Filter’, Astrobiology, 20(5), pp. 572–579. doi: 10.1089/ast.2019.2154.
Kipping, D. (2020) ‘An objective Bayesian analysis of life’s early start and our late arrival’, Proceedings of the National Academy of Sciences, 117(22), pp. 11995–12003. doi: 10.1073/pnas.1921655117.
Schmidt, G. A. and Frank, A. (2019) ‘The Silurian hypothesis: would it be possible to detect an industrial civilization in the geological record?’, International Journal of Astrobiology, 18(2), pp. 142–150. doi: 10.1017/S1473550418000095.
Seckbach, J. (2012) Genesis – In The Beginning, Cellular Origin, Life in Extreme Habitats and Astrobiology, Vol. 22. Edited by J. Seckbach. Dordrecht: Springer Netherlands (Cellular Origin, Life in Extreme Habitats and Astrobiology). doi: 10.1007/978-94-007-2941-4.
Spada, G. and Melini, D. (2020) ‘Evolution of the number of communicative civilizations in the Galaxy: implications on Fermi paradox’, International Journal of Astrobiology, 19(4), pp. 314–319. doi: 10.1017/S1473550420000063.
Webb, S. (2015) If the Universe Is Teeming with Aliens … WHERE IS EVERYBODY?, If the Universe is Teeming with Aliens… Where is Everybody? Cham: Springer International Publishing (Science and Fiction). doi: 10.1007/978-3-319-13236-5.
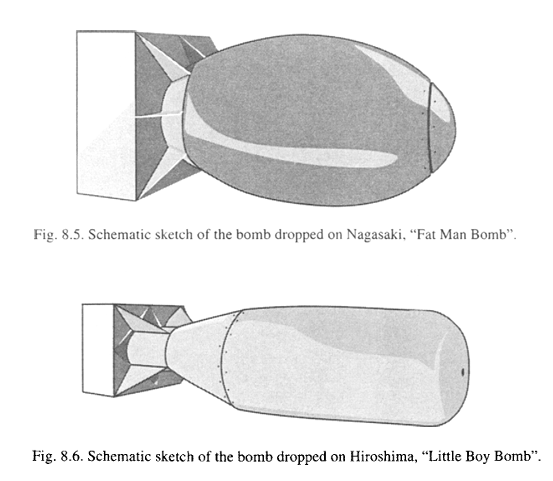
Trinitita: la gema radiactiva de la primera explosión nuclear
escrito por C. Menor-Salvan | 3 octubre, 2023
C. Menor-Salván, 15 de julio 2023.
A pesar de lo que sugiere su nombre, la trinitita no es un mineral. Ni es natural. Es un material que se formó durante un hecho histórico decisivo: La primera explosión nuclear de la Historia, el 16 de Julio de 1945, en Alamogordo (Nuevo México, EEUU). Este artículo es una ampliación del publicado en The Conversation. Añadimos aquí más datos, anécdotas y explicación de algunos conceptos básicos sobre radioisótopos.
Una nueva y temible arma
Se acercaba el final de la Segunda Guerra Mundial. El Proyecto Manhattan llevaba desarrollándose en secreto desde 1941, cuando el presidente Roosevelt, motivado por la posibilidad de que Alemania consiguiera desarrollar un nuevo tipo de arma de destrucción masiva, autorizó la puesta en marcha de uno de los proyectos técnicos y científicos mas ambiciosos de toda la Historia. El proyecto supuso un enorme esfuerzo contrarreloj que implicó a físicos, químicos, ingenieros, matemáticos y miles de trabajadores. Los hitos científicos del proyecto fueron muy importantes y dieron un impulso al desarrollo industrial y tecnológico de los EEUU. También plantó la semilla de la Guerra Fría, debido a su dramático fruto, surgido en julio de 1945 de las instalaciones de Los Álamos, en Nuevo México: The Gadget
‘Ground-zero’: la torre e instalación de la prueba Trinity. La bomba se instalaría en la caseta de la parte superior de la torre. Toda esta instalación se volatilizó literalmente en la explosión.
Las instalaciones y el desarrollo del proyecto eran secretos, pero la Ciencia siempre es abierta, lo cual dio lugar a curiosas anécdotas. En 1943, una de las revistas de Ciencia Ficción más conocidas, Astounding Science Fiction, publicó el relato ‘Deadline’ de Cleve Cartmill, en el que se contaba el desarrollo de una bomba atómica de increíble poder destructivo. Los detalles eran sospechosamente similares a lo que se estaba desarrollando para The Gadget, por lo que el FBI investigó al autor y al editor, John W. Campbell. No encontraron evidencias de una fuga de información y Campbell explicó que, a partir de la información científica disponible, cualquier persona con algo de conocimiento y sentido común puede extrapolarlo a una historia de ciencia ficción realista. Campbell convenció al FBI de que censurar el relato sería más sospechoso que dejarlo tal cual. De hecho, tras la publicación, se recibieron algunas cartas diciendo que la historia era ridícula e imposible. Se cuenta que Campbell dijo a los agentes, «por cierto, ¿están construyendo la bomba en el desierto en Nuevo México, verdad?» Parece ser que varios científicos eran lectores de la revista y cambiaron al mismo tiempo su dirección a un apartado postal en Santa Fe. A pesar del secreto, mucha gente ya sabía que algo importante se estaba construyendo, dados los movimientos inusuales de científicos y los pedidos de productos y materiales. Es imposible mantener en secreto las conspiraciones, en especial si la ciencia está por medio.
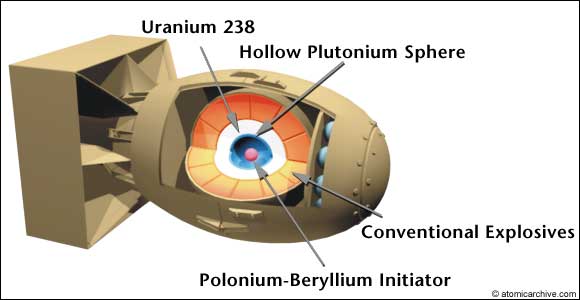
The Gadget fue el primer prototipo funcional de una bomba nuclear. El dia 15 de Julio de 1945, estaba montado y listo para explotar durante el test Trinity, destinado a comprobar experimentalmente las predicciones teóricas. The Gadget era un dispositivo de implosión: un explosivo convencional comprime el núcleo de plutonio-239 de la bomba, que alcanza su masa crítica y provoca una reacción de fisión en cadena que libera una cantidad de energía jamás vista.
El Gadget montado en la caseta sobre la torre de 30 metros, preparado para la prueba
Plutonio-239
El plutonio-239 es un isótopo fácilmente fisible (ver más adelante) fabricado por irradiación de uranio con neutrones. Tiene la ventaja de presentar mayor probabilidad de fisión, requerir menor cantidad El plutonio no se encuentra en la Naturaleza mas que en forma de trazas, en algunos yacimientos de uranio donde se formaron ‘reactores nucleares naturales‘.
Por ello, uno de los retos del proyecto era obtener suficiente plutonio puro a partir de uranio. Una confusión común del público respecto al combustible nuclear es si puede usarse como arma. Los elementos fisibles, como el plutonio-239, para poder usarse como explosivo deben ser muy puros. Muchísimo más que en su uso como combustible nuclear. En los reactores nucleares, la energía producida por la fisión se libera de modo no explosivo, generando mucho calor que se usa para producir electricidad. El plutonio, para utilizarse en una bomba atómica, debe primero fabricarse y luego purificarse cuidadosamente, para que alcance una gran pureza tanto química como isotópica. En resumen, no podemos ‘robar’ combustible nuclear convencional y fabricar una bomba atómica. Haría falta una planta de procesado.
Uranio metálico puro. El uranio se obtiene a partir de sus minerales y se enriquece en uranio-235, fisible, mediante un complejo proceso. El uranio-238, isótopo mayoritario, se transforma en plutonio-239 durante las reacciones nucleares que tienen lugar.
En el proyecto, utilizaron el Reactor B de la planta secreta de producción de plutonio de Hanford (Washington). Ahora museo, entonces fue el primer reactor comercial para enriquecimiento de combustible nuclear, operado por la empresa DuPont; la empresa renunció a los beneficios comerciales, limitándose a cubrir la inversión, y se desmarcó de este área para no ser asociada con el desarrollo de la bomba, pues consideraron que daría mala imagen.
El reactor B era un reactor nuclear con una potencia de 250 MW, pero no destinado a producir electricidad, sino a generar plutonio-239 a partir del uranio-238, por captura neutrónica impulsada por la fisión del uranio-235. La reacción de síntesis de plutonio-239 tiene lugar en varias fases:
1- el uranio-238 captura un neutrón liberado por la fisión del uranio-235. Se transforma en uranio-239.
2- El uranio-239 tiene un periodo de semidesintegración (ver más adelante) de tan sólo 23 minutos. Se transforma por emisión beta (emite electrones, transformándose un neutrón en un protón) en neptunio-239. Este, a su vez, con un periodo de semidesintegración de 2.35 días, emite radiación beta y se transmuta a plutonio-239, que se acumula gracias que tiene un periodo de semidesintegración de más de 24000 años.
El combustible nuclear utilizado contiene hasta un 1.5% de plutonio, siendo mas o menos el 50% plutonio-239; se procesa tras la reacción, en un complejo proceso químico y metalúrgico (llamado método PUREX), para extraer y purificar el plutonio. Actualmente muchos reactores nucleares siguen este principio y, además de producir electricidad, autogeneran más combustible nuclear que permite aprovechar el uranio-238 natural no fisible y que puede usarse en otros reactores. El sueño de los alquimistas de la transmutación de los elementos era posible, aunque no como ellos imaginaban.
El desarrollo del proyecto Manhattan requirió mucha Química. Los análisis de todos los materiales, extracción de plutonio o uranio y la preparación de los diversos componentes llevó al desarrollo de procesos químicos. Todas las técnicas analíticas usadas se recopilaron en este libro.
Un hecho poco conocido por el público es que esta reacción de producción de plutonio-239 fue planeada por el proyecto Tube Alloys, un proyecto de fabricación de un arma nuclear puesto en marcha por el gobierno británico anteriormente al proyecto Manhattan. Debido a su alto coste, el gobierno británico llegó a un acuerdo con el norteamericano, y finalmente se desarrolló en EEUU tras transferir sus avances y predicciones. Si el proyecto Manhattan llegó a su fin fue gracias al esfuerzo de innumerables científicos europeos.
Núcleo del reactor B de Hanford. En él se introducían las barras de combustible nuclear y las barras de control, así como el sistema de refrigeración por agua. La reacción nuclear de fisión del uranio-235 libera neutrones que son capturados por el uranio-238 mayoritario, y que se convierte en plutonio-239.
El concepto de la bomba, desarrollado por el físico Seth Neddermeyer es sencillo (aunque hay detalles complejos, que no voy a discutir): se toma una esfera hueca de plutonio con una masa sub-crítica, es decir, en la que la fisión nuclear no entra en cadena al no tener suficiente masa y densidad. Esta masa es de unos 2.5 a 4.5 kilogramos, del tamaño de una pelota de ping-pong al tamaño de una naranja. La esfera metálica se recubre de un material reflector y amplificador de los neutrones y se inicia usando dos cosas: un iniciador de polonio y berilio que genera neutrones para ‘arrancar’ la reacción y un explosivo convencional dispuesto de tal modo que, al estallar, comprima la esfera de plutonio. Así, aumenta la densidad, entra en la zona crítica y la reacción nuclear de fisión tiene lugar en cadena, auto-amplificándose y liberando una enorme cantidad de energía en menos de un 1 milisegundo. Con este sistema de implosión pueden manejar el plutonio de forma ‘segura’ y detonarlo cuando se considere.
Esquema de las bombas de implosión The Gadget y Fat Man. El sargento Herbert Lehr, en una de las imágenes más famosas de la historia de la bomba. Es el 12 de julio de 1945 y lleva el núcleo de plutonio de la bomba (ver imagen de más arriba) para su instalación. Como se ve, el núcleo de plutonio es bastante pequeño, aunque muy pesado. El núcleo completo del Gadget pesaba 6 kg.
Ahora somos todos unos hijos de puta
A las 5:29 de la mañana del 16 de Julio de 1945, se detonó el Gadget en el inhóspito desierto de la Jornada del Muerto (sugerente nombre que le pusieron los primeros españoles que llegaron allí).
Fue la primera explosión nuclear de la Historia, con un rendimiento aproximado de 19 kilotones. Fue más potente de lo calculado, destruyendo algunos instrumentos científicos ubicados a supuesta distancia segura. Esta potencia explosiva se había calculado con un test previo, el 100 ton test, en el que se detonaron 100 toneladas de TNT. El test Trinity, usando el test previo como referencia, se estimó que era equivalente a unas 19000 toneladas de TNT. A partir de entonces, se toma esta referencia para estimar el rendimiento o potencia explosiva de un arma nuclear.
Imagen de la explosión de Trinity, el 16 de julio de 1945, tomada desde unos 16 km de distancia por el fotógrafo Jack Aeby. Los científicos y la instrumentación para medidas se ubicó a 9 km de distancia.
Tras la explosión, Kenneth Bainbridge, director científico del test Trinity, exclamó: “Ahora somos unos auténticos hijos de puta” Robert Oppenheimer remarcó que esa fue la frase más apropiada que se dijo tras la explosión. En efecto, las dos siguientes bombas, llamadas Little Boy y Fat Man(siendo esta la versión militar de The Gadget) mataron a unas 214000 personas, de las cuales aproximadamente la mitad murieron por las explosiones y el resto debido a la contaminación radiactiva.
Las bombas. Fat Man era la versión práctica de The Gadget. Little Boy era una bomba de uranio-235, con un diseño algo distinto. Esquema de V. Valkovich, Radiactivity in the Environment, 2000.
El fallout, polvo radiactivo depositado tras la explosión, cubrió un radio de unos 30 km.. A pesar de todo lo que hemos visto, tanto el Gadget como las dos siguientes bombas tenían un poder destructivo muy inferior a las armas nucleares actuales. Esta es la base del terror nuclear que marcó la Guerra Fría.
La trinitita, testigo silencioso de la explosión
El plutonio-239 sostiene una reacción, llamada fisión, en la que un neutrón parte un núcleo atómico en fragmentos, liberando, en este caso, una media de 2.88 neutrones, que rompen otros núcleos de plutonio y así sucesivamente. No todos los isótopos de elementos radiactivos son fisibles, es decir, no todos pueden usarse para llevar a cabo esta reacción nuclear. Los dos elementos más apropiados para ello y que se usan en reactores nucleares y en armas nucleares son el uranio-235 y el plutonio-239. El uranio-238, el isótopo de uranio natural más abundante, no puede sostener una reacción de fisión.
Esta reacción puede amplificarse en cadena cuando se supera la masa crítica con plutonio muy puro. En este caso tiene lugar la liberación explosiva de la energía producida por la ruptura de los núcleos. La fisión de los núcleos genera otros elementos químicos. Estos elementos no son iguales que sus versiones (es decir, isótopos) comunes, sino que son muy radiactivos. La fisión del Pu-239 genera más de 100 elementos químicos, en sus versiones (isótopos) radiactivas.
Isótopo
Los elementos químicos naturales tienen una masa atómica promedio. Pero en la mayoría de los casos, están formados por una mezcla de isótopos. Los isótopos poseen el mismo número de protones y electrones (mismo número atómico, Z), pero distinta masa atómica (A), debido a que tienen diferente número de neutrones. Son el mismo elemento, con las mismas propiedades químicas, pero diferentes propiedades nucleares. Hay isótopos estables e isótopos radiactivos o radioisótopos. El caso más sencillo es el del hidrógeno, que tiene tres isótopos: el hidrógeno (A=1), el deuterio (A=2), ambos estables, y el radioisótopo tritio (A=3).
El plutonio tiene seis isótopos principales, desde A=238 hasta A=244. El más estable es el plutonio-244, que tiene un periodo de semidesintegración de 88 millones de años. Seguramente en la Tierra primitiva existió gran cantidad de plutonio como elemento natural, pero tras 4600 millones de años desde su formación, no quedan más que trazas difícilmente detectables en algunos minerales de uranio.
Los neutrones provocan, además, la transformación de los materiales con los que se encuentran en elementos radiactivos. Este proceso se denomina activación neutrónica: los elementos capturan neutrones que cambian el peso atómico del elemento, convirtiéndolo en un isótopo radiactivo.
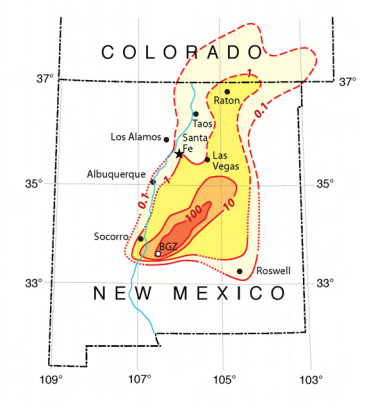
El resultado es, además de la brutal explosión, una contaminación radiactiva que puede alcanzar cientos de kilómetros de distancia, debido a que algunos elementos son volátiles o gases, como el yodo-131 o diversos isótopos de xenón y kriptón. En el test Trinity, se pudo detectar contaminación radiactiva producida por el fallout de la explosión a 150 km de distancia. Pero la contaminación tuvo más alcance. Un empleado de Kodak se dió cuenta de que había contaminación radiactiva gracias a la aparición de ‘puntos’ oscuros en material fotográfico en Indiana, a unos 2000 km del sitio del test. Se realizaaron entonces análisis y se pudo detectar la presencia de cerio-141, otro producto de la fisión del plutonio-239, intensamente radiactivo y con un periodo de semidesintegración de 33 días.
Afortunadamente, la mayor parte de la contaminación radiactiva cayó en zonas no habitadas, y a los pocos días había ya decaído mucho, con la desintegración de los elementos más activos. Sin embargo, aún no está demasiado claro cuánta gente del público quedó expuesta a dosis significativas de radiactividad ni qué ocurrió con el plutonio remanente, esparcido por la zona de la explosión.
Mapa del fallout radiactivo 12 horas tras la explosión del Gadget, medido en mR/h (miliroentgents/hora). Para interpretar el mapa, la isolínea de 0.1 mR/h ya marca un nivel significativo pero aceptable de exposición. La radiación de fondo está entre 0.002 y 0.01 mR/hr y un vuelo comercial produce una tasa de exposición de unos 0.2 mR/h.
La temperatura de la detonación del Gadget superó a la de la superficie del Sol. El calor fundió la arena del desierto levantada por la explosión, que formó gotas de vidrio incandescente que llovieron en un radio de cientos de metros. Cuando todo había terminado, los investigadores vieron que el suelo estaba cubierto por vidrios de colores, normalmente verde, en ocasiones formando bonitas formas transparentes. y que llamaron trinitita. No era anecdótico: se calcula que se formaron unas 1700 toneladas de trinitita, que cubrieron un área de unos 600 metros de diámetro.
Recogieron muestras que se guardaron como recuerdo del hecho histórico. Incluso algunas se usaron para fabricar joyas exclusivas. Pronto se dieron cuenta de que era mala idea. La trinitita contenía elementos producidos por la explosión y era intensamente radiactiva, hasta el punto de provocar quemaduras en la piel. Para evitar accidentes y robos, pues se convirtió en un material preciado por curiosos y coleccionistas, se enterró la mayor parte de la trinitita.
Fragmentos de trinitita, recogidos durante una visita al sitio de la explosión en Alamogordo (Nuevo México, EEUU). Foto: C. Menor-Salván
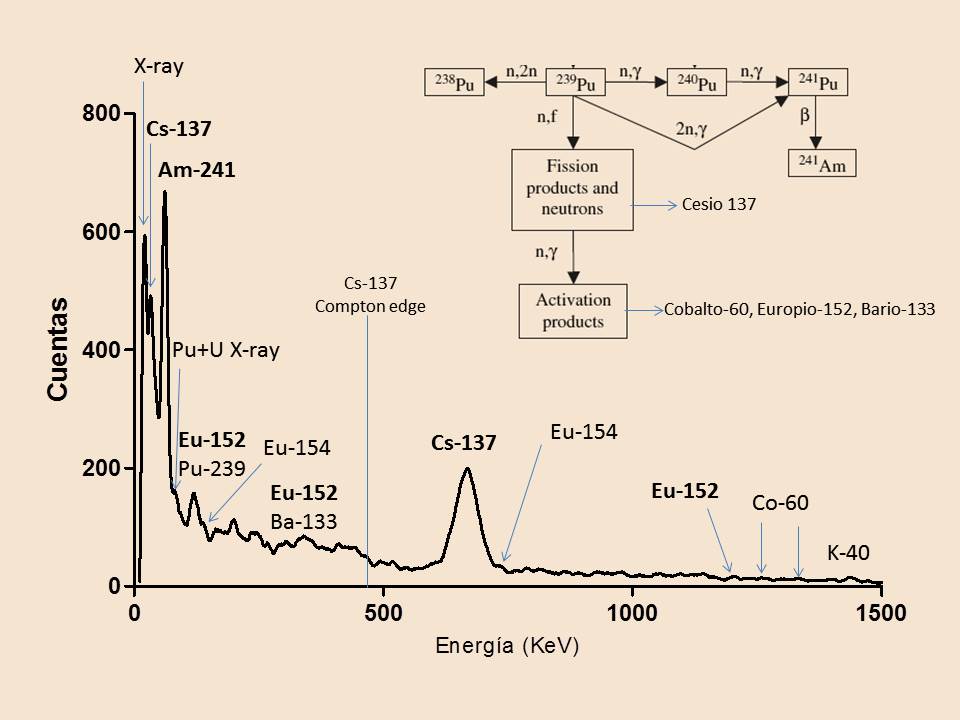
Hoy dia, la trinitita ha perdido la mayor parte de su radiactividad y puede manejarse sin riesgo. Pero aún contiene testigos de la explosión nuclear. En la siguiente figura podéis ver el análisis mediante espectrometría gamma de una muestra de trinitita, realizado en nuestro laboratorio. El espectro resultante que se muestra es de baja resolución, debido a que hemos utilizado un detector de centelleo. La instrumentación para espectrometría gamma de alta resolución utiliza un detector semiconductor. Pero, para nuestro propósito, es suficiente con un análisis a baja resolución.
Espectrometría Gamma
Esta técnica de análisis se basa en la emisión de radiación gamma por los elementos radiactivos. La mayoría de los isótopos radiactivos emiten rayos gamma, fotones de alta energía, como resultado de la relajación de los núcleos atómicos excitados tras las transformaciones nucleares. Cada isótopo radiactivo emite rayos gamma de energía característica. Esta propiedad se puede utilizar para su identificación. Por ejemplo, el cesio-137 emite rayos gamma a una energía característica de 662 KeV (kiloelectronvoltios). Esta emisión gamma se produce al decaer el Cs-137 y transmutarse en Ba-137, emitiendo rayos beta. El núcleo de Ba-137 resultante se encuentra en un estado excitado, metaestable. El exceso de energía se emite como fotones gamma, relajándose a Ba-137 estable.
El espectro gamma se puede registrar utilizando detectores de centelleo, cristales que emiten luz cuando la radiación impacta sobre ellos. Entonces se obtiene un espectro de baja resolución, como el mostrado más arriba. Si se utilizan detectores semiconductores, se obtiene el espectro de alta resolución. Este tipo de detector es mas costoso y requiere enfriamiento, normalmente usando nitrógeno líquido. Por ello, en muchas aplicaciones se siguen usando detectores de centelleo, muchísimo más asequibles.
La espectrometría gamma se puede utilizar para identificar contaminación radiactiva o para analizar materiales mediante activación por neutrones.
El análisis revela los isótopos radiactivos mayoritarios presentes aún en la muestra, tras 78 años desde la explosión:
Cesio-137: es uno de los principales productos de fisión del plutonio. Con un periodo de semidesintegración de 30 años, es el mayor elemento radiactivo de la trinitita. El cesio-137 es uno de los principales causantes de la contaminación radiactiva en una explosión nuclear.
Americio-241: indica que la bomba estaba formada por plutonio-239. Se forma mediante un proceso de captura neutrónica. Un átomo de plutonio-239 captura dos neutrones, bien secuencialmente o de una vez, transformándose en plutonio-241. El Pu-241 tiene un periodo de semidesintegración de 14 años, transformándose en americio-241 mediante emisión de radiación beta. El Am-241 formado tiene un periodo de semidesintegración de 410 años, por lo que será detectable durante siglos. Este elemento es muy conocido, pues forma parte de los antiguos detectores de humo iónicos.
Bario-133: es difícil de detectar, pues su actividad se reduce a la mitad en algo más de 10 años. Se piensa que su origen está en el explosivo que se usó para detonar la bomba, llamado Baratol, formado por nitrato de bario. El bario-133 se forma por captura de neutrones (activación) del bario-132 natural. Aunque el rendimiento es bajo, pues este isótopo
Europio-152: un elemento radiactivo característico de la trinitita, utilizado en la confirmación de ésta frente a falsificaciones. Se piensa que se formó por activación neutrónica del europio natural presente en la arena del desierto.
Cobalto-60: Casi indetectable, debido a que tiene un periodo de semidesintegración de sólo 5 años. Su presencia en la muestra se debe a la activación neutrónica del cobalto de las aleaciones de acero de la torre de prueba y todo el equipamiento.
Periodo de semidesintegración
Este concepto es fundamental en radioquímica. Es el tiempo necesario para que la actividad de una muestra de un isótopo radiactivo se reduzca a la mitad. Por ejemplo, en el caso del americio-241, su periodo de semidesintegración es de 410 años. Esto significa que, en una muestra de americio-241, tienen que pasar 410 años para que la mitad del número total de átomos del isótopo se desintegren y, por tanto, su actividad se reduzca a la mitad. En el caso del bario-133, su periodo de semidesintegración es de 10 años. Esto quiere decir que, en la trinitita, la actividad inicial de bario-133 se ha reducido a la mitad casi 8 veces. Por ello, la actividad de este elemento es ínfima.
No se debe confundir este concepto con el de vida media. Esta es el tiempo promedio necesario para que un átomo dado se desintegre. Es un concepto relacionado, pero no equivalente, ya que el periodo de semidesintegración hace referencia a la población total de átomos, y la vida media es, digamos, la 'esperanza de vida' de un átomo individual. En español se confunden los términos a veces debido a que en inglés, half-life es equivalente a periodo de semidesintegración y, por tanto, half-life no debe traducirse como vida media.
El periodo de semidesintegración es una constante física y no depende de ningún factor externo. Gracias a ello se utiliza en medidas de datación en geociencias, por ejemplo. El decaimiento de elementos radiactivos nos permite determinar la edad de muestras en arqueología, paleontología y geología. Un ejemplo es la prueba del carbono-14, que permiten determinar la edad de un material orgánico en la investigación histórica y arqueológica.
El periodo de semidesintegración nos permite hacernos una idea de la ‘intensidad’ o ‘peligrosidad’ de un isótopo radiactivo. De modo similar a la combustión de una vela: cuanto más rápida, más intensa la luz y mayor es la temperatura, pero menos dura. Cuanto menor es el periodo de semidesintegración, más intensa es la radiación emitida, porque hay un mayor número de átomos desintegrándose al mismo tiempo. Actualmente, en la trinitita permanecen los elementos de mayor periodo de semidesintegración, por ello la radiactividad de ese material ya carece de peligro. En 1945, los isótopos muy radiactivos, de muy bajo periodo de semidesintegración, hacían que la trinitita fuera un material peligroso.
Isótopos radiactivos producidos en la detonación nuclear. La mayoría son de vida muy corta, por lo que la contaminación radiactiva, extremadamente intensa durante los primeros días tras la explosión, decrece rápidamente.
Fragmentos de trinitita
Los elementos radiactivos desaparecerán con el tiempo, pero las características peculiares de este material, la presencia de estructuras extrañas como los cuasicristales, y las huellas isotópicas que delatan su origen, seguirán ahí tras la desaparición de la civilización. Un testigo de nuestro paso por el planeta y del genio, soberbia y maldad humanas.
Referencias
Day, J. M. D., Moynier, F., Meshik, A. P., Pradivtseva, O. V. & Petit, D. R. (2017) ‘Evaporative fractionation of zinc during the first nuclear detonation’, Science Advances, 3(2), p. e1602668. doi: 10.1126/sciadv.1602668.
Parekh, P. P., Semkow, T. M., Torres, M. A., Haines, D. K., Cooper, J. M., Rosenberg, P. M. & Kitto, M. E. (2006) ‘Radioactivity in Trinitite six decades later’, Journal of Environmental Radioactivity, 85(1), pp. 103–120. doi: 10.1016/j.jenvrad.2005.01.017.
Salter, L. P. & Harley, J. H. (1965) ‘“Trinitite”: Cobalt-60, Cesium-137, and Europium-152’, Science, 148(3672), pp. 954–955. doi: 10.1126/science.148.3672.954.
Valković, V. (2019) ‘Radioactive nuclides in nature’, Radioactivity in the Environment, pp. 1–29. doi: 10.1016/b978-0-444-64146-5.00001-x.
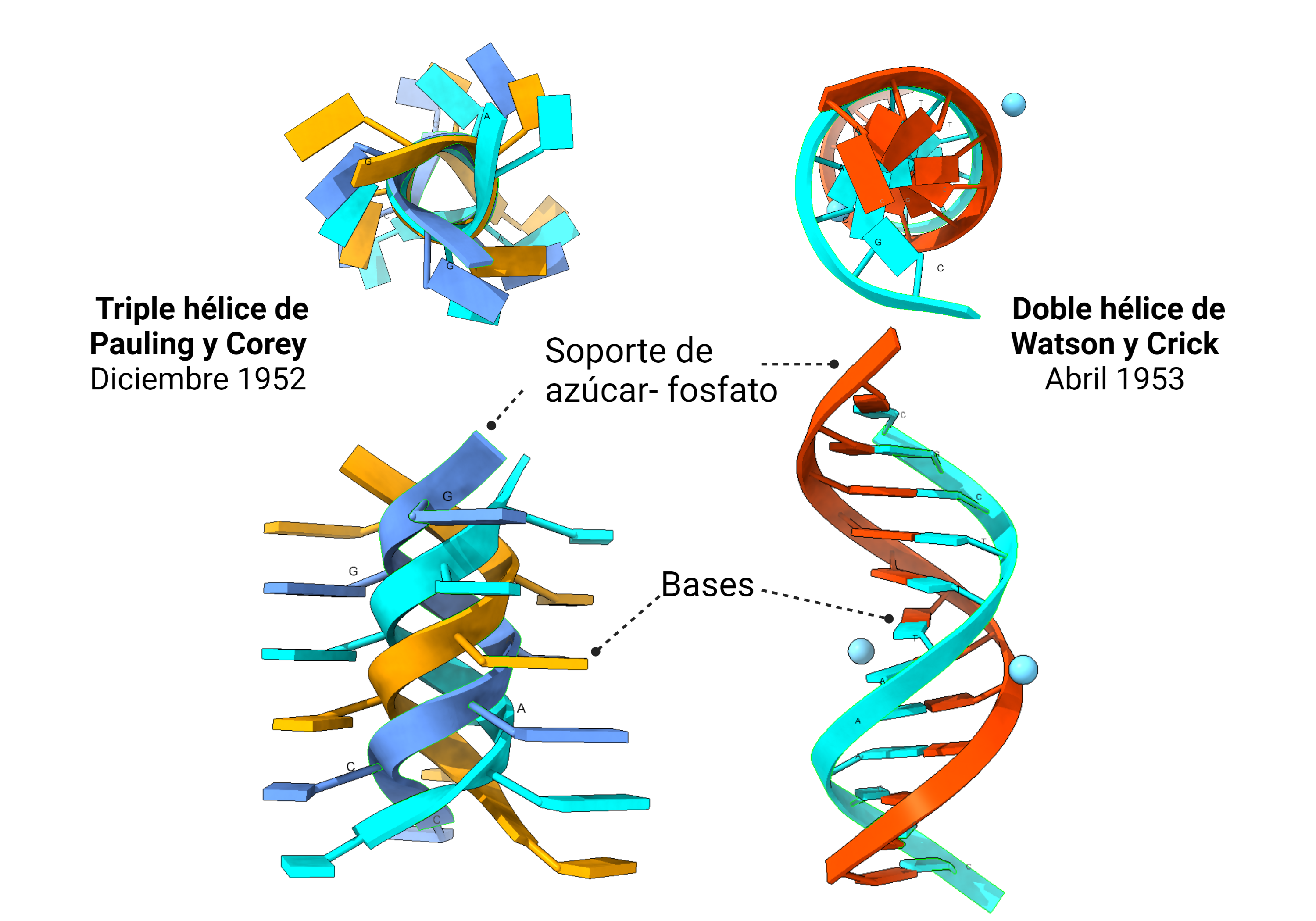
El error de Pauling y la carrera por la estructura del ADN
escrito por C. Menor-Salvan | 3 octubre, 2023
C. Menor-Salván. Ver. 2.5. Abril 2023
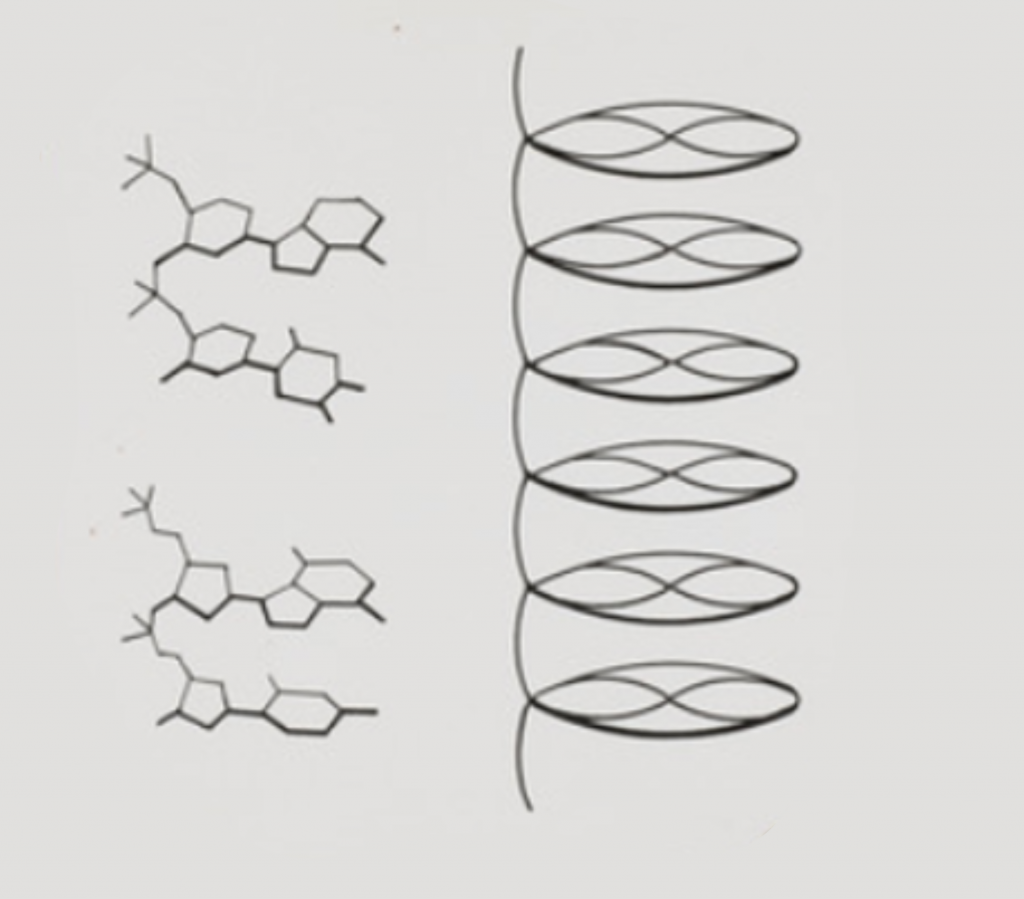
El descubrimiento de la estructura del ADN fue uno de los logros científicos destacados del siglo XX. En él participaron además de los conocidos Watson y Crick y Rosalind Franklin, una serie de científicos relevantes, cuyo nombre apenas se recuerda; sin sus contribuciones, no podemos entender la historia completa. Esta aventura nos enseña además que, en la ciencia, aunque se hagan famosos los «goleadores», el conocimiento se construye de modo colectivo y los errores pueden ser tan importantes como los aciertos. Delicioso es el fruto que surge tras el amargor del error y la ignorancia, y de ellos es desde donde se construye la ciencia. Además, los aspectos mundanos pueden ser tan relevantes como los técnicos.
El ADN se descubrió en el siglo XIX, pero se tardó más de medio siglo en revelar su estructura
No hay que confundir el descubrimiento de la estructura del ADN, con el descubrimiento del ADN en sí. Se atribuye el descubrimiento del ADN al químico suizo Friedrich Miescher, entre 1860 y 1874, siendo Phoebus Levene quien dio, en 1909, su descripción química precisa. Miescher propuso, en 1874, que, de alguna manera la «nucleína» (nombre que dió al ADN) era la «causa específica de la fertilización».
Levene acuñó el término ‘ácido nucleico’. Propuso la ‘teoría del tetranucleótido’, sugiriendo que el ADN estaba compuesto por cuatro bases, un azúcar y fosfato. Sin embargo, en aquel momento, aún no existía la tecnología necesaria para entender la arquitectura de la molécula.
Aspecto real del DNA puro. Esta muestra se obtuvo de timo de vaca, la fuente usual de DNA para su estudio. De hecho, en la época del descubrimiento de su estructura, no se le llamaba DNA sino «ácido timonucleico». Imagen: C. Menor-Salván/UAH
Pasó casi medio siglo hasta que se determinó su estructura. Una vez disponible el arsenal técnico adecuado, no había nadie mejor para lograrlo que el genial químico estadounidense Linus Pauling (1901-1994), quien estuvo a las puertas de conseguirlo antes de que Watson, Crick, Wilkins y Franklin publicaran su famoso artículo triple de abril de 1953.
Pauling fue uno de los científicos más relevantes del siglo XX. Recibió el premio Nobel en Química en 1954 por su contribución al conocimiento de los enlaces químicos. Para esa fecha había realizado tantos descubrimientos importantes en numerosos campos de la Química y la Biología, que, cuando le llamaron para comunicarle la concesión del premio, pidió a su interlocutor que le leyese la comunicación, pues no tenía claro por cuál de sus trabajos recibía el premio.
Pauling había resuelto otro gran problema: la estructura de las proteínas. Con Robert Corey y Herman Branson, publicaron en 1951 las estructuras secundarias que ahora ilustran los libros de texto. Este importante trabajo era ya suficiente para que tuviera fama imperecedera en el mundo de la Ciencia, pero Pauling era ambicioso y estaba obsesionado con resolver todas las estructuras de macromoléculas biológicas. Así, era el favorito en la carrera por el ADN. Él mismo estaba convencido de ello.
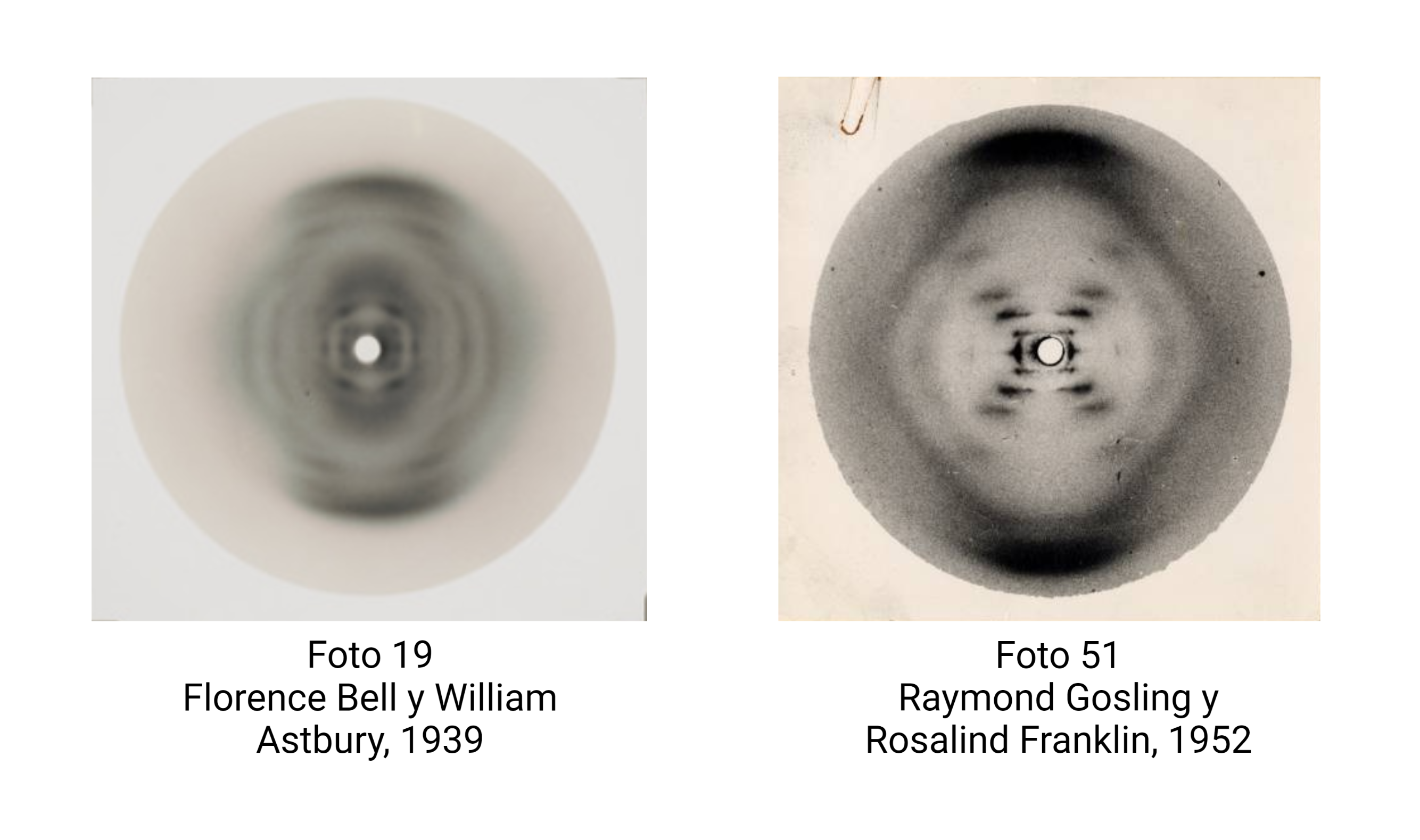
Astbury, Bell y la triple hélice de Pauling
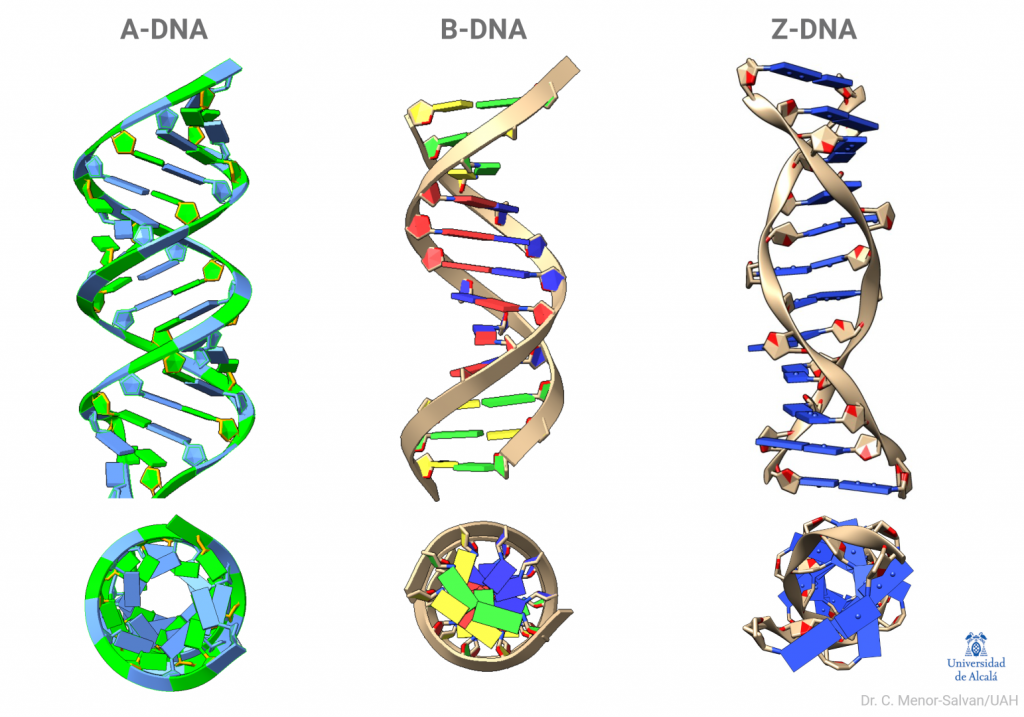
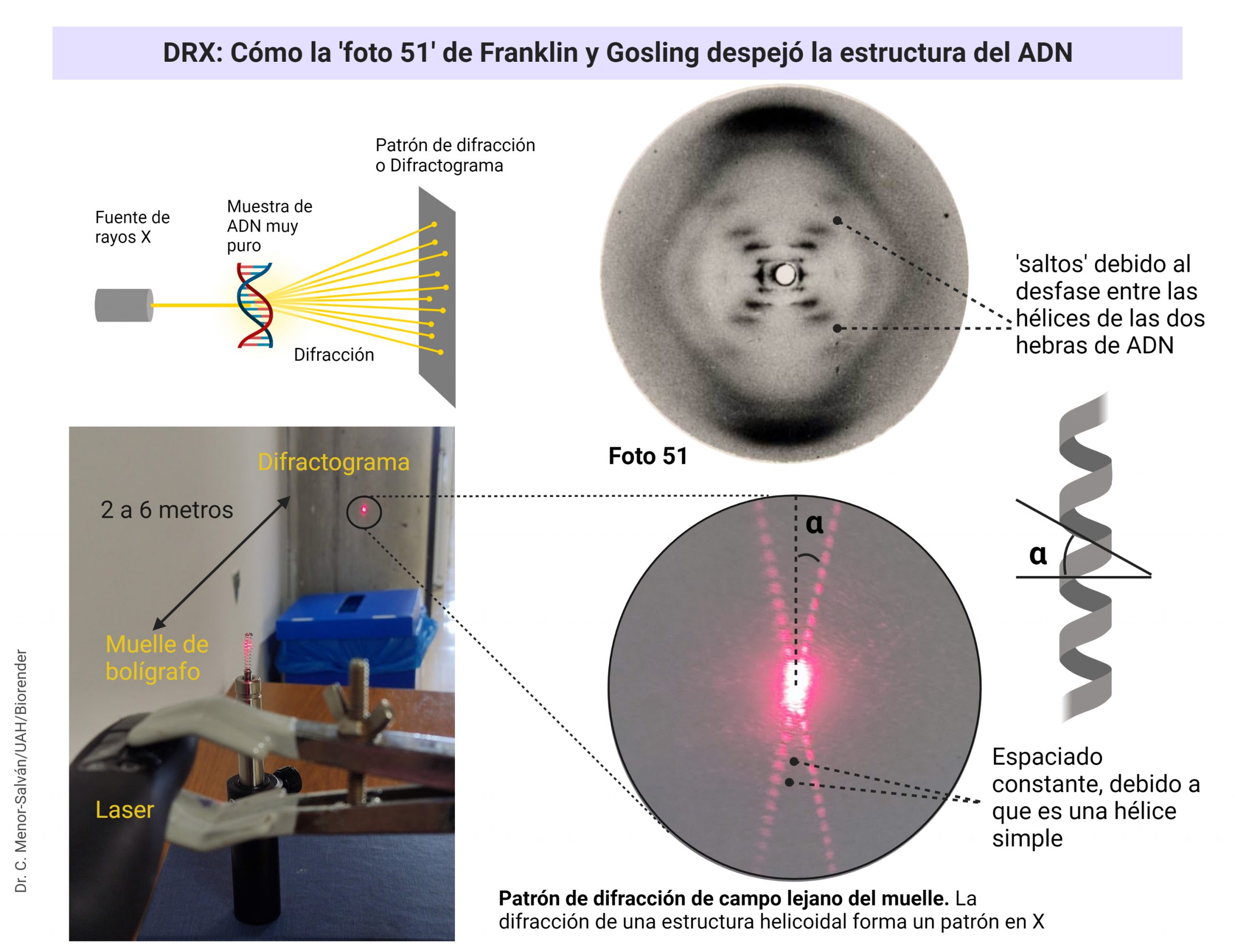
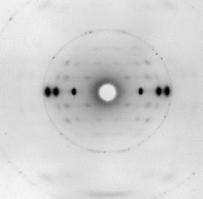
Una brillante química británica, Florence Bell, presentó, en 1939, su tesis sobre la estructura del ADN y las proteínas, bajo la dirección de William Astbury. En ella, Bell y Astbury presentaron, por primera vez en la Historia, imágenes de difracción de rayos X de ADN. Esta compleja técnica, que se aplica en campos desde la Bioquímica Estructural hasta la Mineralogía, es esencial para resolver las estructuras macromoleculares así como las estructuras de los sólidos cristalinos. Y ahí está el problema: EL ADN es muy difícil de cristalizar y tiene una peculiaridad clave, que descubrió Rosalind Franklin: puede cambiar de forma.
Portada de la tesis doctoral de Florence Bell. Su lectura es muy interesante, está redactada con un tono honesto y cálido, y constituye un hermoso documento sobre el nacimiento de la Biología Molecular. Después, con el inicio de la Segunda Guerra Mundial, todo quedó en un impasse; entre tanto, Bell se casó con un oficial norteamericano y emigró a EEUU tras la guerra. A pesar de los esfuerzos de Astbury, Bell no quiso continuar su carrera científica, aunque trabajó como química en EEUU.
Bell logró obtener un esbozo de la estructura del ADN, formado por largas fibras con las bases colocadas en paralelo y unidas, a modo de cuentas de collar, por el fosfato; a partir de ello, no pudo avanzar más. Su tesis relata los intentos para obtener una muestra de ADN lo suficientemente cristalina, que terminaban en imágenes de difracción borrosas. El ADN se resistía a revelar su estructura. Había algo que Bell ignoraba: el ADN no tiene una única forma. Al preparar el ADN puro, se formaba una mezcla de lo que ahora conocemos como ADN B y ADN A, que hacían muy difícil interpretar los datos. Además, todavía tenía que mejorar la tecnología para la obtención de éstos.
Fue Rosalind Franklin, 15 años después, quien logró lo que Bell no pudo: obtener una forma única de ADN cristalino, en su forma canónica ADN B, con el que hizo posible la famosa foto 51, que aclaró definitivamente la estructura.
Florence Bell propuso en su tesis de 1939 este esbozo de la estructura de ADN, en el que los nucleótidos, unidos por enlaces fosfodiester 3′-5′, formaban largas cadenas con las bases apiladas. Si Bell hubiera conseguido mejores datos de DRX, seguramente ella misma, o con Pauling, no habrían tenido ningún problema en determinar la estructura. Imagen: F. Bell, 1938
Aun así, Bell proporcionó datos importantes, gracias a una imagen particularmente clara, la imagen 19, tales como la distancia entre las bases y que los nucleótidos formaban largas cadenas.
Pauling estudió los resultados de Bell y Astbury, tomando como hipótesis que las bases del ADN se orientan al exterior, para enlazarse con otras moléculas y que los fosfatos se organizan en un patrón similar al de algunos minerales. También tuvo en cuenta otro dato fundamental: el biofísico Robley Williams había logrado, en 1952, obtener una imagen en 3D de una molécula de ADN con un microscopio electrónico. Pauling observó que ese pequeño cilindro podría ser una trenza helicoidal.
Con estas ideas, propuso un modelo en triple hélice, en un artículo enviado, no sin prisas, el 31 de diciembre de 1952. En él, publicado en febrero de 1953, Pauling sugiere que su modelo es todavía algo preliminar y requería refinado. Su colaborador Corey le avisó de que había problemas, como que el modelo no encajaba del todo bien, no se podían incluir iones de sodio, a pesar de que el ADN formaba una sal sódica, y estaba el problema de la repulsión de las cargas del fosfato; Pauling reconoció que su modelo estaba algo «apretado». Pero la prisa estaba justificada: sabía que, en Gran Bretaña, Maurice Wilkins y su equipo estaban obteniendo imágenes de difracción del ADN y que unos entonces desconocidos James Watson y Francis Crick estaban trabajando en un modelo de ADN.
Sin embargo, el modelo de Pauling resultó ser erróneo.
Comparación del modelo erróneo de Pauling (izquierda) y el DNA B canónico de Watson y Crick (derecha). La diferencia fundamental es que Pauling preparó un modelo en el que las bases se orientan hacia el exterior. Pauling pensaba que las bases formarían puentes de hidrógeno con otras moléculas, como proteínas. Sin embargo, aunque encajaba más o menos bien con los datos de Bell y Astbury, el modelo tenía problemas muy profundos, como ignorar las cargas de los fosfatos. Imagen: C. Menor-Salván
Pares de bases y la precaución de Wilkins
El modelo en triple hélice encajaba mas o menos bien con los datos de difracción de rayos X de los que disponía Pauling. Es más, el primer modelo de ADN que idearon Watson y Crick, en 1951, era una triple hélice similar a la de Pauling. La diferencia es que ellos contaron con Rosalind Franklin. Cuando mostraron a Franklin su modelo, ella les dio razones convincentes por las que el modelo no era válido. Franklin ya había observado la importancia del agua y de los iones en la estructura (ella estaba trabajando con la sal sódica del ADN) y de cómo debían distribuirse los fosfatos. Ellos la escucharon y volvieron a los cálculos de un nuevo modelo. En ese momento, Franklin y Wilkins, aunque no estaban a favor de determinar un modelo aún, también pensaban que se trataba de una triple hélice. Tendría que llegar la foto 51, obtenida por Franklin, a despejar las dudas: la molécula de ADN era una doble hélice.
Comparación de la foto de DRX de ADN favorita de Bell, la foto 19 (izquierda) con la histórica foto 51 de Franklin y su doctorando Gosling. Franklin realizó un descubrimiento clave: el ADN se presentaba en dos formas, la forma A y la forma B (actualmente sabemos que hay alguna más), mutuamente intercambiables por deshidratación y rehidratación y con diferente grado de coordinación con el agua. Aislar las formas permitió obtener imágenes de DRX claras, algo que Bell no logró, al obtener mezclas de las dos estructuras.
Las claves científicas del error de la triple hélice de Pauling las explica Rosalind Franklin en su magnífico artículo de 1953, en el que muestra que tenía clara la estructura: los datos de Pauling no eran lo suficientemente resolutivos; además, una hélice con los fosfatos en el interior y las bases en el exterior no encaja, ni con las propiedades del ADN, ni con la química del fosfato; y, no menos importante, Pauling no tuvo en cuenta un hallazgo previo fundamental al que Franklin si da crédito.
En 1947, un joven bioquímico, Michael Creeth, propuso un modelo de ADN formado por dos cadenas unidas por puentes de hidrógeno entre sus bases. Los mentores de Creeth eran Gulland y Jordan, quienes habían demostrado el apareamiento entre bases del ADN. Esta observación, para Franklin, tal como ella misma cuenta en su artículo de 1953, es fundamental. Considerando los pares de bases y los datos de difracción, la famosa doble hélice emergía como la estructura correcta.
Modelo propuesto por Michael Creeth para la estructura del ADN, tal como figura expuesto en el National Centre for Macromolecular Hydrodynamic, en Nottingham (Inglaterra). Actualmente, Bell y Creeth, así como sus mentores, son grandes olvidados en el descubrimiento del ADN.
Consciente de que los datos de Bell y Astbury no tenían resolución suficiente, Pauling escribió a Wilkins, solicitando ver los suyos. Wilkins, que no deseaba que el gran Pauling tomara el control de la investigación, se negó. Suele decirse que Pauling no tuvo oportunidad de reunirse con ellos directamente, debido a que el gobierno de EEUU le denegó la renovación del pasaporte para viajar a Gran Bretaña en 1951. Lo cierto es que el impacto de esta anécdota no fue grande, pues, finalmente, Pauling visitó Inglaterra durante un mes en 1952, coincidiendo con el trabajo clave de Franklin y, durante el cual, lamentablemente, no prestó atención al ADN.
Mientras Pauling, pensando que Wilkins seguiría negándose a colaborar, se centraba en sus trabajos con las proteínas, Franklin y su doctorando Gosling obtenían las imágenes históricas que confirmaron la estructura del ADN. Franklin no tenía inconvenientes para mostrar sus resultados y, si Pauling hubiera hablado con ella directamente, la historia del ADN sería distinta. Es más, ocurrió algo extraño: Franklin mostró sus imágenes a Corey, mano derecha de Pauling. Corey sugirió a Pauling que el modelo de triple hélice no encajaba bien y que había varios problemas. Pero él insistió, pensando que, bueno, ya resolverían los problemas pendientes. No visitar a Franklin y no escuchar a Corey fue un error histórico.
No fue el único error de Pauling. Watson y Crick tuvieron en cuenta otro dato clave: el bioquímico austriaco Erwin Chargaff les explicó que las bases del ADN siguen una proporción muy sencilla, que hoy conocemos como reglas de Chargaff. Estas apoyaban la idea de que las dos cadenas estarían unidas por las bases. Pauling y Chargaff se conocieron en 1947 durante un crucero de vuelta a EEUU y hablaron de ello; pero, considerándole un tipo «molesto y desagradable», Pauling despreció sus observaciones.
El modelo erróneo de Pauling fue el impulso definitivo que llevó a Watson y Crick a publicar, sólo dos meses después, su doble hélice. Cuando el artículo de Pauling apareció en febrero de 1953, los británicos estaban sorprendidos: era un modelo ingenuo, erróneo, similar al que ellos concibieron en 1951 y que Franklin les hizo desechar, y que, como los mismos Watson y Crick comentaron, incluso violaba las reglas químicas básicas del fosfato, expuestas por el propio Pauling en su famoso libro de texto de Química General. Eufóricos por la oportunidad que el error de Pauling les brindaba, era el momento perfecto para arrebatarle la gloria de resolver la estructura del ADN; se apresuraron, entonces, a publicar su modelo antes de que Pauling tuviera tiempo de darse cuenta de su fallo y rehacer su modelo, a la vista de los nuevos datos de difracción de rayos X.
Página del famoso artículo de Watson y Crick, en la que figura su modelo de doble hélice y agradecen específicamente a Wilkins y Franklin por sus contribuciones. Watson y Crick eran teóricos y dependían de los datos experimentales. La foto que se ve en la parte inferior no es de Franklin, sino de Wilkins.
Así, en abril de 1953, tras un acuerdo con Wilkins y Franklin, se publicaron tres artículos: uno por Watson y Crick, otro por Wilkins y el tercero por Franklin y Gosling, detallando la estructura del ADN y los datos esenciales que la sostienen. Sorprende que mucha gente piense que aquel mes se publicó un sólo artículo, firmado por Watson, Crick y Wilkins, y que Franklin fue ignorada completamente. Como ocurre con El Quijote, que todo el mundo lo conoce, pero casi nadie lo ha leído. Sin embargo, la lectura de los artículos aclara cosas, como que, en efecto, no fue un artículo, sino tres, cada uno preparado por los líderes del descubrimiento: Watson y Crick eran los teóricos que desarrollaron el modelo, y Wilkins y Franklin los experimentalistas que lograron preparar el ADN puro y obtener los datos de su estructura. A priori no parece mal arreglo. Con ojos actuales, cada uno de ellos tendría su paper histórico en Nature como IP (Los problemas de índole personal que llevaron a que Wilkins y Franklin publicaran por separado, o por qué publicaron tres papers separados en lugar de uno todos juntos, no son objeto de esta entrada, y es un tema que se discute mucho, aunque para mi forma parte del gossip científico, no de la ciencia en sí)
Aceptando la derrota
Pauling aceptó sus errores con elegancia y, en la Conferencia Solvay de ese mismo abril de 1953, expresó su apoyo al modelo de Watson y Crick:
«Aunque solo han pasado dos meses desde que el profesor Corey y yo publicamos nuestra estructura propuesta para el ácido nucleico, debemos admitir que probablemente esté equivocada; Aunque se podría hacer algo de refinamiento, creo que es muy probable que la estructura de Watson-Crick sea esencialmente correcta»
En 1988, durante una conversación informal en un congreso, Pauling recapitulaba:
«Supongo que siempre pensé que la estructura del ADN era mía para resolver y, por lo tanto, no la perseguí con suficiente agresividad».
Definitivamente, no escuchar las señales de que su modelo no era correcto y su ambición, que le llevó a estar convencido de que era el único que podría resolverla, no le ayudaron.
Anexo 1: Cómo ayudó la foto 51 a revelar la estructura
La técnica de difracción de rayos X (DRX) es y ha sido esencial para la Biología. Es una técnica compleja, costosa y que requiere de una larga formación específica, por lo que los cristalógrafos, como lo fueron Rosalind Franklin, Florence Bell o el propio Pauling, expertos en DRX, son muy valiosos para la investigación y su labor no siempre es suficientemente reconocida. Prácticamente todas las estructuras que podemos encontrar en el Protein Data Bank, por ejemplo, han sido resueltas mediante DRX.
Para obtener un buen patrón de difracción, o difractograma, es necesario disponer de una muestra muy pura del compuesto que se está analizando, uniforme y cristalina (es decir, todas las moléculas de la muestra están ordenadas). Esto fue muy difícil de conseguir con el ADN, en especial por su tendencia a la transición entre ADN A y ADN B según su contenido en agua.
El DNA no tiene una única estructura. Son posibles múltiples conformaciones.
Rosalind Franklin y Raymond Gosling (a quien nadie recuerda, a pesar de haber trabajado en el laboratorio codo con codo con Franklin) consiguieron una imagen excepcional, como hemos visto: la famosa foto 51. Esta foto se reproduce en muchas ocasiones, pero ¿qué significa? ¿cómo se interpreta?
Usando un simple muelle y un puntero laser para entender el patrón de difracción de una hélice
La interpretación completa es muy compleja, pero podemos entenderla de modo sencillo, en especial si usamos un experimento: si tomamos un muelle de boli o cualquier otro muelle pequeño y fino y lo iluminamos con un láser, que tenga el haz cuanto más grueso mejor (de modo que ilumine varias vueltas de hélice al mismo tiempo), el muelle va a difractar la luz laser. Como la diferencia entre la distancia de vueltas del muelle y la longitud de onda del laser es muy alta, el patrón de difracción que se produce es la difracción de campo lejano o difracción de Fraunhofer, que podemos ver al proyectar la luz a una distancia de entre 2 y 6 metros del muelle. El patrón de difracción nos permite medir con mucha precisión las medidas del muelle. Si cambiamos el muelle por moléculas ordenadas de un compuesto, y el láser por un haz fino de rayos X, muy penetrante y de longitud de onda muy pequeña, comparable a las dimensiones de la molécula, obtendremos un patrón de difracción de su estructura.
El patrón de difracción de una estructura en hélice da lugar a una «X» muy característica. El ángulo de la «X» y la distancia entre las manchas luminosas nos permite calcular las medidas de la hélice (ver mas abajo). Con el sencillo experimento del muelle y el puntero láser, podemos reproducir el experimento y los cálculos que realizaron Franklin y Gosling.
Si nos fijamos en las imágenes de Florence Bell, este patrón no está nada claro, debido a las dificultades que tuvo para preparar la muestra. Pauling supo que la molécula de ADN era helicoidal porque vio una imagen de microscopía electrónica.
Otro aspecto clave de la foto 51 es que revelaba claramente que el ADN no estaba formado por una hélice, sino por dos hélices unidas, con un desfase entre ellas. Este desfase daba lugar a los huecos que se pueden ver en la foto. Este dato confirmaba definitivamente que el ADN estaba formado por dos hebras unidas formando una hélice doble. Franklin pudo medir la molécula usando el patrón, obteniendo un ajuste muy bueno con el modelo teórico propuesto por Watson y Crick.
¿Qué habría ocurrido si el modelo de Creeth hubiera sido correcto? El patrón de difracción habría sido completamente distinto. Podemos modelizarlo también mediante la difracción de Fraunhofer de dos alambres paralelos muy juntos:
Esto nos da un patrón de difracción o difractograma lineal:
Como en el caso anterior, con el espaciado entre las manchas luminosas podemos medir la distancia entre los dos alambres. Comparemos este difractograma con el difractograma de rayos X real de un poliester, un polímero lineal formado por cadenas paralelas:
Y aquí vemos el patrón de difracción de un fragmento proteico formado por láminas beta paralelas:
Así queda demostrada la estructura de la molécula usando un método físico clave, la DRX. La verdad es que las cosas son como tienen que ser: si el DNA tuviera la estructura de Creeth, la Biología Molecular sería imposible, y por tanto la vida. Pero eso todavía no lo sabían en los años 1940-1950.
Anexo 2: Extracto de los artículos históricos de abril de 1953
Watson y Crick mencionan el modelo erróneo de PaulingÚltimo párrafo del artículo de Rosalind Franklin de abril de 1953, relatando sus observaciones y concluyendo que «la idea general no es incosistente con el modelo propuesto por Watson y Crick». Como experimentalista, apoya con sus datos el modelo teórico propuesto. Franklin, además, cita el trabajo clave de Gulland y sus colaboradores, descubridores del apareamiento de bases en el ADN. El modelo de Pauling fue el ‘punching ball‘ de los tres artículos. Aquí, Franklin discute por qué el modelo de Pauling es erróneo. El modelo de Pauling, a pesar de ello, fue un catalizador de la publicación de estos tres trabajos históricos. El resto del artículo de Franklin es un relato detallado de los datos e interpretación de los análisis por difracción de rayos X. Primer párrafo del trabajo de Franklin. Aquí relata algo esencial que, a veces, se pasa por alto: el descubrimiento de las formas B y A del ADN. Franklin reconoce aquí a otro científico realmente olvidado por la Historia del descubrimiento del ADN: el noruego Sven Furberg, quien, en 1948, estuvo también a punto de descubrir la estructura del ADN y propuso, por primera vez, que debía tener una estructura helicoidal. Pero Furberg tropezó en la misma piedra que Bell y Astbury…
Referencias
ASTBURY, W. T. & BELL, F. O. (1938) ‘X-Ray Study of Thymonucleic Acid’, Nature, 141(3573), pp. 747–748. doi: 10.1038/141747b0.
Dahm, R. (2008) ‘Discovering DNA: Friedrich Miescher and the early years of nucleic acid research’, Human Genetics, 122(6), pp. 565–581. doi: 10.1007/s00439-007-0433-0.
FRANKLIN, R. E. & GOSLING, R. G. (1953) ‘Molecular Configuration in Sodium Thymonucleate’, Nature, 171(4356), pp. 740–741. doi: 10.1038/171740a0.
Gann, A. & Witkowski, J. A. (2013) ‘DNA: Archives reveal Nobel nominations’, Nature, 496(7446), p. 434. doi: 10.1038/496434a.
Harding, S. E., Channell, G. & Phillips-Jones, M. K. (2018) ‘The discovery of hydrogen bonds in DNA and a re-evaluation of the 1948 Creeth two-chain model for its structure’, Biochemical Society Transactions, 46(5), pp. 1171–1182. doi: 10.1042/BST20180158.
Lake, J. (2001) ‘Why Pauling didn’t solve the structure of DNA’, Nature, 409(6820), pp. 558–558. doi: 10.1038/35054717.
Levene, P. a, Hydrolysis, A. & Rockejeller, T. (1917) ‘Yeast Nucleic’, J. Biol. Chem., 40, pp. 415–424.
Pauling, L. & Corey, R. B. (1953) ‘A Proposed Structure For The Nucleic Acids’, Proceedings of the National Academy of Sciences, 39(2), pp. 84–97. doi: 10.1073/pnas.39.2.84.
Pederson, T. (2020) ‘The double helix: “Photo 51” revisited’, The FASEB Journal, 34(2), pp. 1923–1927. doi: 10.1096/fj.202000119.
Ronwin, E. (1953) ‘The Phospho-Di-Anhydride Formula and Its Relation to the General Structure of the Nucleic Acids’, Science, 118(3071), pp. 560–561. doi: 10.1126/science.118.3071.560.
Schmidt, G. (1950) ‘NUCLEIC ACIDS, PURINES, AND PYRIMIDINES’, Annu. Rev. Biochem, 19, pp. 149–186.
Thompson, J. et al. (2018) ‘Rosalind Franklin’s X-ray photo of DNA as an undergraduate optical diffraction experiment’, American Journal of Physics, 86(2), pp. 95–104. doi: 10.1119/1.5020051.
WATSON, J. D. & CRICK, F. H. C. (1953) ‘Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid’, Nature, 171(4356), pp. 737–738. doi: 10.1038/171737a0.
El plegamiento de proteínas y las mutaciones del código genético
escrito por C. Menor-Salvan | 3 octubre, 2023
C. Menor-Salván ,oct 2022. V2 Nov 2023
En el año 1969 ya se conocía mucho sobre la estructura de las proteínas y hacía 10 años que había nacido la Biología Molecular moderna. Ya se entendía la relación entre el código genético, la secuencia de aminoácidos de un péptido y la proteína final. Poco tiempo antes, el bioquímico Christian Anfinsen había descubierto que una proteína desnaturalizada podía recuperar su estructura y actividad en cuestión de milisegundos, por lo que quedaba demostrado que la estructura estaba, de alguna manera, codificada en su secuencia.
Para Cyrus Levinthal esto era paradógico: una secuencia de aminoácidos tiene un número enorme de posibilidades estructurales, por lo que las diferentes conformaciones posibles no pueden tener la misma probabilidad y el plegamiento no es resultado de la búsqueda de la conformación nativa, sino que debía ser dirigido por un camino específico y predeterminado. La resolución de la paradoja de Levinthal, es decir, entender cómo se pliegan las proteínas, es uno de los problemas más complejos de la Bioquímica y la Biofísica, y sólo ahora, gracias a la inteligencia artificial y a los métodos computacionales modernos, estamos en condiciones de resolverla completamente.
La paradoja de Levinthal: si un péptido de 100 aminoácidos puede generar tantas conformaciones, más que átomos hay en el Universo, ¿cómo es posible que se pliegue formando proteínas bien definidas?. A la derecha, Cyrus Levinthal con unos estereomicroscopios clásicos de Bausch&Lomb.
El plegamiento proteico es un problema muy complejo, incluso para las células que generan las proteínas: los fallos o, simplemente, las diferentes soluciones al plegamiento estable de un péptido están detrás de enfermedades como el Alzheimer ,las enfermedades por priones o las amiloidosis. Pero, aunque sea tan complejo, podemos señalar algun aspecto interesante, como es el papel esencial de los aminoácidos hidrofóbicos y su relación con la evolución del código genético.
Los inicios de la aplicación de los métodos computacionales a la estructura de proteínas. Actualmente, un PC portátil es miles de veces más potente que esas enormes computadoras. Aun así, fueron claves para la comprensión de las primeras estructuras proteicas, como la de la mioglobina o la lisozima. Este sistema, llamado ‘Kluge’ fue utilizado por Levinthal.
El colapso hidrofóbico
El proceso de plegamiento proteico es gradual, formándose regiones, llamadas foldones, que van adquiriendo conformaciones similares a la forma nativa final. Estos foldones son cooperativos y, cuando empiezan a formarse, favorecen el plegamiento del resto de la proteína. Esto se denomina la hipótesis del foldón.
En la formación de las regiones plegadas es esencial la estabilización energética que proporciona el efecto hidrofóbico, es decir, la expulsión del agua y el secuestro de los grupos R de los aminoácidos hidrofóbicos en el interior de la estructura plegada, dejando los grupos polares y cargados expuestos al solvente. Este efecto es uno de los responsables de la extraordinaria capacidad de las enzimas como catalizadores: al tener un núcleo hidrofóbico, la exclusión del agua y la desolvatación de los sustratos de las enzimas promueven la actividad enzimática.
Esta fase de plegamiento de la proteína, en la que adquiere su estructura nativa en regiones y se ha producido el colapso hidrofóbico, es muy dinámica: la estructura se mueve, los foldones se forman y reconvierten de un modo fluido. Este estado se denomina glóbulo fundido. Finalmente, la proteína alcanza un estado similar a un sólido tridimensional y se estabiliza, formando el estado nativo de la proteína. En este estado, aparecen otras interacciones, como los puentes disulfuro, el stacking de residuos de aminoácidos aromáticos y la unión de cofactores, que terminan de ‘fijar’ la estructura nativa.
En estas condiciones, tenemos la proteína plegada y funcional. Así puede cristalizar, lo cual es muy importante para conocer la estructura. Pero el estado nativo no es totalmente sólido: la proteína puede desnaturalizarse, o volver al estado de glóbulo fundido y cambiar sus conformaciones en respuesta a algún estímulo.
Estructura de la lisozima (derecha), mostrando su plegamiento nativo, en el que se observan dos regiones, una con hélices alfa y, en la parte inferior, un motivo de lámina beta antiparalela. En amarillo, los puentes disulfuro que estabilizan la estructura. Izquierda: Estructura mostrando los residuos hidrófobos, que ‘colapsan’ en el núcleo de la proteína, creando un centro donde el agua queda excluida. Centro: Estructura mostrando los resíduos polares y con carga, que se organizan hacia el exterior, en contacto con el agua o con otras estructuras. Cristal de lisozima, la proteína mostrada en la imagen anterior. La cristalización de las proteínas es un proceso en muchos casos necesario para poder determinar la estructura tridimensional de las proteínas. Los cristales obtenidos, como éste, generado en nuestro laboratorio, permiten obtener las coordenadas de los átomos de la estructura mediante difracción de rayos X.
La idea del colapso hidrofóbico puede parecer contraintuitiva: como es posible que una fuerza débil, como la fuerza de Van der Waals entre grupos apolares, sea lo que dirige el plegamiento de la proteína. ¿cual es el papel de los enlaces de hidrógeno?. Al principio se pensaba que los enlaces de hidrógeno, mucho mas fuertes, eran esenciales en la estabilización de la estructura proteica. Actualmente sabemos que los enlaces de hidrógeno juegan un papel director durante el proceso de plegamiento, ya que dirigen la formación de diversas estructuras secundarias, que se agrupan en motivos y van dando lugar al proceso cooperativo de plegamiento. Sin embargo, no sólo no son esenciales en la estabilización de la estructura nativa, sino que, en ocasiones, son desestabilizadores que favorecen los cambios de conformación. La clave no es la fuerza de las interacciones hidrofóbicas per se, sino el efecto termodinámico, como la variación de entropía asociada al efecto hidrofóbico, que favorece el proceso.
Dada la importancia del colapso hidrofóbico, está claro que los aminoácidos hidrofóbicos van a ser claves en la evolución de las estructuras. Ello ha condicionado la evolución del código genético.
Estructura de la mioglobina, una de las primeras proteínas cuya estructura terciaria fue resuelta. A la derecha, aminoácidos hidrófobos, orientados hacia el interior de la estructura. A la izquierda, aminoácidos cargados, orientados hacia el exterior de la estructura. Algunos forman puentes salinos, que estabilizan la estructura y, en el caso de la mioglobina, juegan un papel importante relacionado con su función. En rojo, grupo hemo. Si rotamos la molécula y la observamos longitudinalmente, se ve mejor el efecto. A la izquierda, aminoácidos hidrófobos Leu, Ile y Val, orientados hacia el interior. A la derecha, aminoácidos cargados Asp, Glu y Lys orientados al exterior.
Código genético, mutaciones y la preservación de la estructura proteica
La secuencia de bases del ADN sufre constantes cambios: mutaciones que cambian bases en la secuencia y que se traducen en cambios en la secuencia de una proteína. Estos cambios pueden ser deletéreos, pero también silenciosos o incluso beneficiosos, dando lugar a nuevas funciones. El código genético ha evolucionado de tal forma que las mutaciones mas comunes, llamadas mutaciones missense, en las que un par de bases sustituye a otro, limiten sus efectos sobre las proteínas resultantes. Afortunadamente, el código genético, que se compone de unas ‘palabras’ de tres letras, llamadas codones, esta degenerado. Esto quiere decir que los 64 posibles codones (‘palabras’ de tres letras formadas con las bases A, G, C y U, es decir 43) van a codificar para tan solo 20 aminoácidos distintos. Ello implica que algunos aminoácidos van a tener hasta 6 codones sinónimos. Esto es posible gracias al balanceo o wobble del RNA de transferencia: la tercera posición del anticodon puede reconocer diferentes bases en el codón, gracias a la formación de pares de bases no Watson-Crick, o la presencia de bases no canónicas en esa posición.
Estructura terciaria de un RNA de transferencia, mostrando sus características más importantes: el patrón de minihélices que se repite, la terminación CCA en 3′ con el aminoácido unido en su posición, y el anticodón, cuya base en la posición 34 es la posición de balanceo o wobble y puede puede formar puentes de hidrógeno alternativos. Por ejemplo, una uridina en el anticodón puede reconocer tanto una adenina como una guanina en el codón.
Gracias al balanceo, hay tan solo 32 diferentes tRNA para todos los codones, y estos transportan sólo 20 aminoácidos (es decir, hay más de un tRNA por aminoácido en algunos casos). Por ejemplo, un tRNAarg de las levaduras tiene el anticodón GCI (donde I es inosina, una base que usualmente no está presente en el DNA ni el RNA). La posición I es de balanceo y puede unirse a los codones CGA, CGU y CGC. Ahora, imagina que hay una mutación y un codón CGU se transforma en un codón CGC. Esta sería una mutación silenciosa, pues, al traducirse el gen, seguiría codificando para una arginina y no se producirían cambios en la proteína. La degeneración del código genético hace posible que cambie el aminoácido sólo en un 25% de las mutaciones de este tipo. Ello aporta estabilidad al genoma: durante la evolución, solo perduraron los genomas capaces de mantener sus estructuras funcionales en un ambiente con muchos cambios. Así, la degeneración del código genético y la limitación del número de aminoácidos pudo aportar una ventaja selectiva a los organismos, reduciendo los efectos de las mutaciones. Un potencial organismo con otro código que implicara más aminoácidos debió ser inviable con las mismas tasas de mutación.
Vista simplificada del efecto de las mutaciones según la posición del codón. Las mutaciones representadas estan basadas en un modelo real, la evolución del SARS-CoV-2 hacia nuevas variantes. El cambio de asparagina a tirosina representa el caso habitual: no cambia la clase de aminoacido (ambos son aminoacidos polares, y la estructura global se va a mantener) pero las propiedades son lo suficientemente distintas como para que se produzcan cambios funcionales o de estabilidad en la proteína. En este caso, la tirosina tiene un anillo aromático, lo que le permite crear interacciones acidicionales.
Aun así, el efecto del wobble no es suficiente; es necesario proteger la función de las proteínas, y, para ello, es clave mantener su estructura. Así, los aminoácidos hidrofóbicos están especialmente protegidos, debido a su paper fundamental en el plegamiento proteico. Cuando se produce una mutación en la primera posición del codón, se va a producir un cambio de aminoácido. Pero, en general, el cambio da lugar a la sustitución de un aminoácido por otro de propiedades similares. Por ejemplo, la valina, un aminoácido hidrófobo, esta codificada por un codón GUU. Si hay una mutación en la tercera posición y se transforma en GUC, sigue codificando para valina, y estamos ante una mutación silenciosa. Pero si hay una mutación en la primera posición y se convierte en AUU, el aminoácido cambia a isoleucina, que también es hidrófobo y tiene propiedades similares. Si la mutación lo convierte en CUU, el aminoácido cambia a leucina, que, de nuevo, es hidrófobo y tiene propiedades similares. Estos cambios no alteran significativamente el plegamiento proteico, ya que el colapso hidrófobo va a seguir sucediendo, por lo que, a pesar de las mutaciones, el plegamiento (y por tanto, la función) se va a mantener.
El problema surge cuando se produce un cambio de un aminoácido a otro de un tipo diferente. Esto puede provocar un cambio más o menos drástico en la estructura o propiedades de la proteína. Es el caso, por ejemplo, de la anemia falciforme, en la que hay una mutación de una base en el gen de la cadena beta de la hemoglobina. El aminoácido mutado cambia el codón para un ácido glutámico CTC por el codón para la valina CAC.
Mutación en la cadena beta de la hemoglobina, que provoca un cambio de un aminoácido con carga a un aminoácido hidrofóbico.
El cambio elimina un aminoácido cargado, el glutámico, por un aminoácido hidrófobo, la valina. Al desaparecer la carga, desaparece la repulsión electrostática entre unidades de hemoglobina, y se sustituye por un aminoácido hidrófobo superficial, que provoca una interacción hidrófoba entre cadenas. Como resultado, la hemoglobina precipita en forma de fibras. De nuevo, los aminoácidos hidrófobos son claves en la estructura. Aquí, la introducción del aminoácido hidrófobo valina tiene consecuencias relevantes, provocando la enfermedad de la anemia falciforme.
Aunque un cambio de aminoácido no tenga gran efecto estructural, influyen en la variabilidad genética y pueden tener importancia. Los SNP o polimorfismos de nucleótido único, son cambios en la secuencia de genes que se traducen en modificaciones de algún aminoácido en las proteínas. Estos SNP crean muchas variaciones entre individuos de la misma especie, y, normalmente, no tienen ningún efecto.
Sin embargo, en algunos casos, los cambios afectan a aminoácidos que alteran ligeramente la función de la proteína (haciendo una enzima mas activa o menos activa, o modificando la afinidad de una proteína por un sustrato). Aunque un cambio de afinidad sea pequeño, tienen una gran importancia farmacológica, explicando la diferente respuesta a algunos fármacos entre individuos. Entre diversas especies, los cambios son mayores para la misma proteína (o para la proteína ortóloga: proteína que ejerce la misma función en otro organismo y proviene del mismo ancestro común, y que normalmente son estructuralmente equivalentes). Esto permite trazar la evolución de las especies y de las propias proteínas.
Referencias
Alas-Guardado, S. de J., Rojo, A. and Merino, G. (2011) ‘La paradoja de Levinthal: cuando una contradicción se vuelve lógica’, Educación Química, 22(1), pp. 51–54. doi: 10.1016/S0187-893X(18)30114-9.
Baldwin, R. L. & Rose, G. D. (2013) ‘Molten globules, entropy-driven conformational change and protein folding’, Current Opinion in Structural Biology, 23(1), pp. 4–10. doi: 10.1016/j.sbi.2012.11.004.
Braakman, R. & Smith, E. (2013) ‘The compositional and evolutionary logic of metabolism’, Physical Biology, 10(1). doi: 10.1088/1478-3975/10/1/011001.
Englander, S. W. & Mayne, L. (2014) ‘The nature of protein folding pathways’, Proceedings of the National Academy of Sciences of the United States of America, 111(45), pp. 15873–15880. doi: 10.1073/pnas.1411798111.
Francoeur, E. (2002). Cyrus Levinthal, the Kluge and the origins of interactive molecular graphics. Endeavour, 26(4), 127–131. doi:10.1016/s0160-9327(02)01468-0
Honig, B. (1999) ‘Protein folding: From the levinthal paradox to structure prediction’, Journal of Molecular Biology, 293(2), pp. 283–293. doi: 10.1006/jmbi.1999.3006.
Karplus, M. (1997) ‘The Levinthal paradox: yesterday and today’, Folding and Design, 2, pp. S69–S75. doi: 10.1016/S1359-0278(97)00067-9.
Kurata, S., Weixlbaumer, A., Ohtsuki, T., Shimazaki, T., Wada, T., Kirino, Y., Takai, K., Watanabe, K., Ramakrishnan, V. & Suzuki, T. (2008) ‘Modified uridines with C5-methylene substituents at the first position of the tRNA anticodon stabilize U·G wobble pairing during decoding’, Journal of Biological Chemistry, 283(27), pp. 18801–18811. doi: 10.1074/jbc.M800233200.
Pak, D., Root-Bernstein, R. & Burton, Z. F. (2017) ‘tRNA structure and evolution and standardization to the three nucleotide genetic code’, Transcription, 8(4), pp. 205–219. doi: 10.1080/21541264.2017.1318811.
Root-Bernstein, R., Kim, Y., Sanjay, A. & Burton, Z. F. (2016) ‘tRNA evolution from the proto-tRNA minihelix world’, Transcription, 7(5), pp. 153–163. doi: 10.1080/21541264.2016.1235527.
¿Por qué el número de la suerte es el 7?. Sobre dados, entropía, vida y orden.
escrito por C. Menor-Salvan | 3 octubre, 2023
C. Menor-Salván. UAH. Actualizado Octubre 2023
El concepto de entropía y su relación con la Bioquímica es uno de los más complicados de entender por parte de los estudiantes. En este artículo, vamos a tratar, sin entrar en formalismos matemáticos, de aclarar el concepto de entropía, el significado del Segundo Principio de la Termodinámica y la relación con la vida y el metabolismo.
Introducción: Entropía NO es desorden
El concepto de entropía, una de las funciones de estado termodinámicas (es decir, una función que describe las características térmicas y energéticas de un sistema), es uno de los que más dificultades presenta para su comprensión. En mi enfoque de la enseñanza de la Bioquímica, la entropía tiene un papel esencial, pues es una de las claves que explican la propia vida y sus estructuras. La entropía es esencial para entender el funcionamiento de las enzimas y para entender por qué el metabolismo no es una colección de reacciones diseñada para fastidiar a los estudiantes, sino que tiene una lógica comprensible.
Una de las razones por las que los estudiantes tienen dificultades con el concepto de entropía es debido a una simplificación confusa que se transmite constantemente en la enseñanza: que la entropía es una medida del desorden. Mayor desorden=mayor entropía. En un texto de 2º de Bachillerato he podido leer:
La entropía S es una magnitud que mide el grado de desorden de un sistema físico o químico
Esta definición es errónea. Además suele acompañarse con imágenes como esta:
Ilustrando la supuesta relación entre entropía y desorden. Este tipo de imágenes DEBEN EVITARSE al introducir la entropía (en esta imagen, podríamos decir que la imagen de la derecha tiene mayor entropía, pero no por la razón que imaginas)
¿cómo puedo explicar a los estudiantes que la entropía es generadora de orden, y que el orden o estructura biológica es resultado del incremento de entropía, si previamente les hemos engañado con una simplificación errónea?.
Tratando de hacerlo lo más simple y conceptual posible y sin entrar en las matemáticas de la Termodinámica formal, vamos a buscar alternativas que nos permitan acercarnos al significado de la entropía y a entender por qué la entropía genera el orden biológico. Ello es necesario para entender, a su vez, por qué el metabolismo es tal como es.
Dados y estados: buscando otras analogías para entender la entropía
¿sabes por qué el 7 es considerado un número de suerte?. ¿si lanzas DOS dados, crees que hay la misma probabilidad de que salga cualquiera de los resultados posibles (la suma de lo que sale en cada dado)?. Hagamos el experimento: si lanzamos dos dados, el resultado puede ser desde 2 hasta 12. Si los lanzamos muchas veces (he estado una tarde de sábado haciéndolo), obtendremos una distribución como esta:
Distribución de frecuencias en el lanzamiento de dos dados de seis caras. Resultado experimental
Según esta distribución, el 7 es el número mas probable. La razón de la preferencia por el 7 en el lanzamiento de dos dados es, en realidad, bastante obvia. Podemos usarla como una analogía del concepto de entropía: la probabilidad de obtener el número 7 es la mayor, por lo que el número 7 es el estado de mayor entropía. 2 y 12 representan los estados de mínima entropía, razón por la cual son las apuestas mas arriesgadas. Antes de explicar por qué es así, vamos a dejarnos enseñar por el físico aleman Arnold Sommerfeld.
¿por qué encendemos una estufa en invierno [igual este invierno lo vamos a tener difícil…]. Una persona común dirá: para que la habitación esté mas caliente; un físico se expresará hablando de suministrar energía. En este caso, tendrá razón el profano y no el físico
Según Sommerfeld, este ejemplo lo proponía el físico suizo Robert Emden. El caso es que, en efecto, encendemos la estufa para que la habitación esté caliente. Este calor se disipa constantemente a través de las ventanas y las paredes; por ello si apagamos la estufa, la temperatura comienza a disminuir. Aquí no os estoy descubriendo nada nuevo.
Como la habitación es mayoritariamente aire, sabemos que la energía contenida va a depender de la temperatura. Esto se expresa a través de esta ecuación:
Esta ecuación nos dice que la energía U contenida en la habitación es proporcional al número de átomos que hay en la habitación, N, y a la temperatura de la habitación, T. Parece lógico: cuanta más temperatura, más energía.
También sabemos que, si calentamos un gas, va a sufrir o un aumento de presión o una expansión. Como en la habitación la presión va a ser constante, mantener la temperatura constante implica que vamos a mantener constante la energía contenida en la habitación. Pero, mientras la estamos calentando, el aire se va a expandir, saliendo por las rendijas e intercambiándose con el exterior.
¿qué ocurriría si la habitación estuviera perfectamente aislada? Lógicamente, al no poder expandirse, aumentaría la presión y la temperatura. Pero, una habitación normal es un sistemaabierto, es decir, puede intercambiar energía con el exterior, y también materia, pues hay intercambio de aire a través de rendijas. Por lo que el aislamiento perfecto no va a existir.
Rudolf Clausius nos explicó que la variación de entropía de un sistema es el cociente entre el calor suministrado dividido entre la temperatura. Es decir, la entropía no es una medida de desorden, sino una medida de la relación entre energía suministrada y temperatura.
Como queremos mantener nuestra habitación a temperatura y presión constante (no queremos morir, y hay rendijas, por lo que el aire puede expandirse), hemos deducido, a partir de la ecuación anterior y de las propiedades de los gases, que a presión constante, la variación de entropía en nuestra habitación es:
Es decir, que la variación de entropía que tenemos en la habitación es proporcional al incremento de la temperatura desde A a B, y al incremento de volumen del aire desde A a B. Si aumentamos la temperatura, entonces, aumentamos la entropía. n es el número de moles de aire, y cv es el calor específico. Idealmente, una vez que hemos alcanzado una temperatura constante, ya no hay aumento de entropía. Y por tanto, no sería necesario suministrar más calor. Observad que no hablo de orden. El incremento de entropía va asociado al incremento de temperatura y/o volumen. Las moléculas del aire ¿están igual de ordenadas o más ordenadas?. No lo se. Pero si sabemos que, si aumentamos el volumen, por ejemplo, al calentar, les estamos dando a nuestras moléculas más libertad para moverse. Vamos a volver a esta idea de ‘libertad’ más adelante.
Pero, en nuestra habitación ocurre un proceso irreversible: el calor se disipa al exterior de dos formas: irradiándose a través de los materiales y las ventanas, y mediante intercambio de aire caliente del interior con aire frío del exterior.
El famoso dibujo de I. Prigogine, que ilustra la el Segundo Principio de la Termodinámica aplicado a sistemas abiertos. El sistema (habitación), produce entropía y ésta siempre va a ser positiva. O, como mucho, va a ser cero. Pero, hay un proceso de disipación a través de las paredes, y asociado a ese proceso hay una variación de entropía también.
El intercambio de materia y energía de la habitación (nuestro sistema) con sus alrededores, implica que tenemos que descomponer la entropía en dos términos: ΔiS es la entropía generada en el interior de la habitación, que es siempre cero o positiva; ΔeS es la entropía del proceso de intercambio con el exterior y puede ser negativa, cero o positiva. Como la entropía es el cociente entre calor y temperatura, y queremos mantener la temperatura y presión constante en la habitación, entonces:
La temperatura del exterior de la habitación Te es menor que la temperatura del interior Ti. Por tanto, si perdemos todo el calor, o sea, Qi=Qe, entonces el término de entropía de intercambio con el exterior es mayor en valor absoluto. Como el intercambio con el exterior está sacando calor de la habitación, entonces, el término es negativo. Por tanto, la variación global ΔS=ΔiS–ΔeS<0. Hemos visto antes que la entropía depende de la temperatura, por lo que si sacamos el mismo calor que metemos, la habitación se enfría hasta que Ti=Te. Esto es lo que ocurre en una habitación cualquiera: en el momento que apagamos la calefacción, la habitación se enfría. Al final, Qi se igualará con Qe. La clave para mantener la habitación caliente es suministrar energía constantemente y llegar a un estado estacionario en el que Qi>Qe durante un lapso de tiempo.
Total, que para mantener la temperatura constante, y, por tanto, ΔS=0, necesitamos que Qi>Qe, es decir, suministramos más energía de la que perdemos durante el tiempo que queremos estar calientes. Lógico, ¿no?. Por ello nos interesa minimizar Qe, utilizando aislamientos, de modo que podamos minimizar Qi, y reducir la factura de la luz. Aquí voy a añadir una nota: estamos simplificando mucho la discusión, pues nos estamos centrando en el aire, un gas, y además lo consideramos un gas ideal. Cuando hablemos de reacciones químicas, cambios de fase, etc. la discusión se complica, y hablaremos de energía libre, entalpía y entropía.
El término ΔeS negativo lo denominó Schrödingerentropía negativa o neguentropía: la entropía asociada al proceso de disipación. Es decir, estamos neutralizando la acumulación de entropía en la habitación al calentarla, sacándola fuera y aumentando la entropía del exterior.
Pérdidas de calor de una casa. Estas pérdidas representan la entropía negativa: aumentan la entropía del exterior, disminuyendo la entropía del interior.
Esto es lo que define el SEGUNDO PRINCIPIO DE LA TERMODINÁMICA: la entropía total del universo (nuestra habitación + el exterior) siempre aumenta. Por ello, para poder mantener la entropía de la habitación constante (que mantenga su temperatura y presión constantes) necesitamos estar constantemente generando calor en ella y aumentar la entropía del universo.
Nuestra habitación (sistema, puede ser el salon de casa o una célula) y el exterior. Dentro de la habitación aumentamos la entropía (enciendiendo el radiador por ejemplo). Las condiciones de contorno controlan el flujo de entropía con el exterior. Si ambos se igualan, la presión y temperatura de la habitación se mantendrán constantes. Eso requiere un gasto de energía.
Con esto hemos explicado físicamente lo que todos sabemos por experiencia: que si apago el radiador, la habitación se enfría. Por este principio, los ingenieros han hecho enormes esfuerzos en reducir los procesos irreversibles en máquinas y edificios, tales como reducir la disipación de calor mediante aislamiento, disminuir el rozamiento y desgaste en los motores, etc. En la vida real, como vamos a tener procesos irreversibles, siempre tenemos que consumir energía para ‘pagar’ esta especie de impuesto entrópico. Esta es la razón por la que no existe el móvil perpetuo y por la que, hagamos lo que hagamos, siempre se va a enfriar la casa al apagar la calefacción.
Aplicación en la bioquímica
La discusión anterior se puede aplicar exactamente igual a un organismo vivo, ya sea una célula o un animal.
En 1992 se publicó un interesante experimento: los investigadores midieron, usando un calorímetro, la producción de entropía de cerditos en una granja y observaron que la producción de entropía iba creciendo durante el periodo infantil, hasta alcanzar un máximo. Cuando el cerdito entraba en edad adulta y cesaba su desarrollo, la producción de entropía disminuía, e iba disminuyendo gradualmente hasta la vejez.
Es decir, mientras estaba en proceso de morfogénesis activa, generando estructura biológica, la disipación de entropía crecía. En la edad adulta entra en estado estacionario.
La ecuación que vimos antes:
la podemos expresar así:
En la que la velocidad o la variación de la producción de entropía es el balance entre la producción de entropía interna del sistema (es decir, debida al metabolismo y procesos vivos del cerdito) y la disipación de energía al entorno en forma de calor (Q) que es una función que varía con el tiempo (estamos en un sistema dinámico), en relación con la temperatura del sistema. Aquí, el calor Q no hay que confundirlo con temperatura. El calor engloba la irradiación de energía y la disipación de energía transportada por moléculas, como el agua, CO2 y otras moléculas emitidas por el organismo, que ‘esparcen’ o ‘dispersan’ la energía. En estado estacionario, ambos términos se igualan.
Por el contrario, si la tasa de producción interna de entropía es menor que la disipación de calor, la variación de entropía es negativa. Tenemos la ‘neguentropía’ y, en el proceso, es cuando se produce la auto-organización de las estructuras de los organismos vivos.
Ilya Prigogine entra en escena: explicando la vida con el teorema de mínima entropía, sistemas no lineales y estructuras disipativas.
Resulta que estamos disminuyendo la entropía de nuestro sistema, mediante la disipación de energía y el aumento de entropía del exterior del sistema. Hay varios tipos de este sistema peculiar que, espontáneamente, se ha alejado del equilibrio.
Uno de estos sistemas es el metabolismo.
El metabolismo es un conjunto conectado y coordinado de reacciones químicas que permite, según el teorema de mínima entropía de Prigogine, minimizar la entropía producida durante la disipación de energía libre, maximizando la negentropía. Ello lleva a la reducción de entropía interna del sistema y la aparición de estructuras emergentes. Este tipo de sistemas se denominan estructuras disipativas (mas adelante volveremos a este concepto).
El metabolismo permite que los organismos vivos se ‘alimenten de entropía negativa‘ (como expresó Schrödinger). Este concepto aún está en discusión y también se ha sugerido que la evolución marca un camino hacia la maximización de la producción de entropía, provocando un aumento de complejidad que aumente la neguentropía para mantener la estabilidad dinámica. En fin, aquí entramos en un área muy compleja. Tanto que lo llamamos la Ciencia de la Complejidad. Pero como sumario, podemos decir que hay una diferencia fundamental entre un sistema vivo y una máquina: en las máquinas, los ingenieros quieren reducir los procesos irreversibles para aumentar su eficiencia disminuyendo la entropía. En la vida, en cambio, DEPENDE de los procesos irreversibles y la producción de entropía es la que hace que funcione. ¿por qué depende de ellos? porque la vida surge a costa de alejarse del equilibrio y de aumentar la disipación de entropía (el segundo término en la última ecuación)
Un ejemplo relativamente simple de estructura disipativa, muy similar al metabolismo, es la reaccion de Belousov-Zhabotinsky:
La aparición espontánea de patrones durante la reacción, como los de la imagen, que son zonas con diferentes parámetros físicos y químicos, es resultado de la disminución de entropía, mediante la transformación de la energía contenida en el ‘combustible’ de la reacción y el aumento de entropía del exterior. Es una estructura disipativa que depende del aumento de entropía del exterior. Por ello, una estructura disipativa, tal como el metabolismo, sólo puede producirse en un sistema abierto. En un sistema cerrado en el que no pueda intercambiar materia y energía con el exterior, el sistema ‘muere’ rápidamente.
¿qué tienen en común la reacción de Belousov con el metabolismo? Para que un sistema químico tenga estas propiedades, deben establecerse una serie de ciclos y reacciones encadenadas, en los que se alternen reacciones químicas reversibles e irreversibles y se produzca un fenómeno de autocatálisis o de catálisis recíproca. Aquí ya entramos en una complejidad excesiva (alguien pillará el chiste), por lo que lo dejamos en éste punto. Pero, la idea es que para que se de la vida tal como la conocemos, el metabolismo tiene que ser tal como es, y debe contener un sistema de reacciones como la glucólisis, que hacen posible que la disipación de entropía permita formar las estructuras biológicas.
Alguien podría preguntar: entonces, termodinámicamente, ¿la reacción de Belousov es vida?. No. La diferencia fundamental es que un organismo vivo es capaz de regular internamente su comportamiento estructural. La estructura en la reacción de Belousov es resultado de los cambios e influencia de su entorno, sin control interno. Este control estructural y regulación interno, que está relacionado con la información biológica, es lo que llamamos biología molecular. Así, podemos establecer un criterio de demarcación entre la biología molecular y la bioquímica, si queremos.
Reacción de Belousov-Zhabotinsky: un modelo del metabolismo.
Vamos a ver de nuevo el ejemplo de la reacción de Belousov:
Belousov descubrió, en 1951, su famosa reacción tratando de encontrar un modelo de laboratorio para el metabolismo central (glucolisis+ciclo de Krebs). Lo que encontró fué fascinante: una reacción que no era ni un equilibrio ni irreversible, sino que oscilaba entre el estado inicial y el final. Si la reacción se lleva a cabo sin agitación y en una capa fina, tiene lugar la formación de bellas estructuras llamadas patrones de Turing.
Belousov estudió exhaustivamente la reacción y propuso un esbozo de su mecanismo (que tardaría décadas en resolverse); envió sus resultados y los detalles experimentales para ser publicados en una importante revista científica, en 1951. Los revisores recomendaron contra su publicación y el editor respondió a Belousov que su “supuesto descubrimiento” era “imposible”. Seguramente, la idea de que una mezcla química se comportase como un péndulo, o que en un líquido homogéneo surgieran estructuras de composición química diferenciada, violando en apariencia el Segundo Principio de la Termodinámica, pesaba más que el repetir la simple receta de la reacción y verlo con sus propios ojos. El editor dijo a Belousov que tenía que aportar más pruebas. Boris entonces volvió al laboratorio y tras otros seis años de trabajo (la Ciencia requiere tiempo…), envió un nuevo artículo, más elaborado y con un estudio profundo del mecanismo. La respuesta que recibió fue de rechazo y un escepticismo aún mayor. Ni los revisores ni los editores se molestaron en repetir por ellos mismos una reacción extremadamente sencilla de reproducir. Simplemente se dejaron llevar por sus prejuicios. Belousov, que ya tenía 64 años, estaba tan enfurecido y decepcionado que decidió no volver a publicar ningún artículo jamás ni relacionarse con otros científicos. Publicó su receta de la reacción en un simposium, quedando solo un oscuro resumen en ruso en el fondo de algunas bibliotecas.
Cuando Zhabotinsky tomó el testigo, la receta de la reacción de Belousov era conocida en diversos centros en la URSS, aunque se desconocía su autor y origen. Zhabotinsky comenzó, como tema de tesis, a estudiar esa curiosa reacción que circulaba por ahí. En los años 60, ya había realizado un extenso estudio y había descubierto al autor de la reacción original: Boris Belousov. Informó a éste de su trabajo y progresos y Boris le remitió, muy agradecido por recuperar su vieja receta, su manuscrito rechazado original y otro material. Cuando la reacción empezó a ser conocida fuera de la URSS, a finales de los 60 y principios de los 70, aún era conocida como la “reacción de Zhabotinsky”, aunque por el esfuerzo de éste, el crédito para Belousov fué finalmente reconocido. Zhabotinsky mantenía informado de sus progresos a Belousov, pero éste siempre rechazó un encuentro personal, así como asistir al simposio sobre reacciones oscilantes que se había organizado en Praga en 1968. Así, Zhabotinsky y Belousov nunca se conocieron personalmente. Belousov murió en 1970, sin llegar a conocer las implicaciones que tendría su trabajo ni disfrutar del reconocimiento que actualmente se le brinda.
Desde el punto de vista de la termodinámica, podemos usar la reacción de Belousov para entender cómo funciona un sistema metabólico:
En primer lugar no es una sola reacción, sino que se genera un sistema multicomponente, abierto y genera una red o ciclos de reacciones. Uno de estos ciclos es un ciclo autocatalítico en el que los productos son, a su vez, reactivos. Estas características las comparten tanto un sistema metabólico como la reacción de Belousov.
Los patrones de Turing son impredecibles, jamás son exactamente iguales y su estructura depende de pequeñas diferencias en las condiciones iniciales. Son el resultado del acoplamiento entre las reacciones químicas y los procesos de difusión. Esta relación es no lineal y da lugar a múltiples estados estacionarios, igualmente probables. La formación de las estructuras, que manifiestan los estados estacionarios, depende de las fluctuaciones microscópicas. Prigogine lo llamaba «orden a través de la fluctuación». En un sistema biológico, las fluctuaciones son cambios ambientales, epigenética o variaciones genéticas, y las estructuras son el fenotipo.
El sistema (ya sea la reacción de Belousov o el metabolismo) alcanza un estado alejado del equilibrio, en el que maximiza la disipación de energía libre de los reactivos, minimiza la producción de entropía y maximiza la disipación de entropía fuera del sistema, de modo que el sistema se sostiene con la neguentropía y, aunque la entropía global aumenta, dentro del sistema se produce una reducción temporal de entropía. En el vídeo podéis ver uno de los modos de disipación de entropía: las burbujas de CO2, el residuo que produce la reacción, y que se eliminan. Este es el principio de mínima producción de entropía de Prigogine. El sistema evoluciona reduciendo su producción interna de entropía y maximizando la disipación. Pero, la reacción de Belousov no es un buen sistema abierto, y termina acumulando demasiada entropía y «muriendo»
Estado final de la reacción de Belousov: se ha disipado la energía libre, la variación de entropía es cero (está en equilibrio) y se destruyen las estructuras. La composición es homogénea y la energía libre final es inferior a la inicial.
Otro modelo, el modelo de máxima disipación de entropía, sugiere el sistema tenderá al estado estacionario en el que la producción de entropía sea máxima. Es decir, la evolución tiende a maximizar el aumento de entropía del entorno, lo cual puede hacerse mediante el balance entre la producción interna de entropía (que siempre aumenta) y el intercambio con el exterior.
La reacción de Belousov o la vida misma no son más que disipadores de energía libre ygeneradores de entropía. Esto permite la evolución compleja de estructuras durante el proceso de disipación de la energía libre (nutrientes) hacia los resíduos (CO2). Prigogine las llamó estructuras disipativas.
Esto nos lleva al concepto de línea del tiempo: el proceso no se puede revertir. Percibimos el tiempo en cuanto partimos de un estado inicial de baja entropía y/o alta energía libre, para terminar en un estado que ha maximizado la entropía y ha disipado la energía libre en forma de calor. Esta es la razón por la que no se puede viajar al pasado, pues ello violaría el segundo principio de la Termodinámica; viajar al pasado implicaría moverse hacia una reducción de entropía del universo.
La vida surge, como una perturbación temporal, en el proceso de disipación de la energía y del aumento de la entropía del universo. Por ello, la vida y estructuras complejas no surgen en los estados de mínima entropía o de máxima entropía, sino de modo dinámico durante el proceso de disipación de energía y aumento de la entropía del universo.
Otro aspecto interesante de estos sistemas es la adaptación y auto-reparación. Las estructuras disipativas son estables a perturbaciones y pueden absorber éstas y ‘auto-sanarse’. La capacidad de autoreparación se puede observar en la reacción de Belousov si se introducen perturbaciones:
Vida y disipación de entropía en el planeta Tierra
Hay una idea interesante al considerar la vida globalmente: la vida surge en la Tierra como una consecuencia del teorema de la maximización de la entropía. La Tierra recibe energía del Sol y debe disiparla. Si no se disipase, el planeta se habría calentado hasta fundirse. En el proceso de disipación, surge la vida como una estructura que contribuye a mejorar esa disipación (es decir, el segundo término de la última ecuación que escribimos).
La Tierra, globalmente, constituye una gran estructura disipativa en la que:
donde T1 es la temperatura de la superficie del Sol (aproximadamente 6000 K) y T2 es la temperatura de equilibrio del planeta, que es a grosso modo la temperatura a la que la Tierra irradia energía, y que es aproximadamente 300 K. Dado que la energía no se acumula en la Tierra (si así fuera, el planeta se calentaría gradualmente), el segundo término de la ecuación es aproximadamente 3×1024/300 cal/grad g
La Tierra recibe mucha energía y poca entropía, es decir, energía de alta ‘calidad’, y se disipa al espacio con un flujo de entropía mucho más elevado. Ello es lo que asegura que se sostenga la vida en la Tierra. Sin embargo, esta cuenta global no es suficiente para entender cómo surgió y cómo funciona la autoorganización molecular que da la vida.
Pero, en términos globales, la biosfera funciona como ciclos de Morowitz (ver más adelante) en los que los organismos fotosintéticos capturan energía solar, la convierten en hidrógeno y genera un gradiente de potencial, que se disipa, impulsando la generación de biomasa. La energía del hidrógeno se almacena y transporta en forma de azúcares, que son utilizados por otros organismos para reconvertirlos en hidrógeno y generar un gradiente, que impulsa a su vez la generación de biomasa o la producción de entropía en estado estacionario. En cada paso, la energía solar se ‘degrada’, o se ‘esparce’, creando un eficiente flujo de entropía hacia el exterior.
La vida en el planeta no es más que otra estructura asociada al principio de máxima producción de entropía. En un planeta como la Tierra, los flujos de fluidos y energía (Sol, calor interno del planeta) han provocado la autoorganización de estructuras (incluyendo la vida) que maximizan el flujo de entropía. Esto no ocurre en todos los planetas. En el Sistema Solar, el principio de máxima producción de entropía se aplica además en Marte, Titán, Europa o Encelado. Por ello, son lugares de interés astrobiológico.
Obsérvese que en ningún momento hemos relacionado la entropía con una medida del orden a pesar de haber estado un rato hablando de entropía. Hemos hablado de la aparición de estructuras (como es la estructura celular), y del macroestado, es decir, el balance global de calor y la temperatura de la habitación, pero no hemos hablado de ‘orden’. Ahora ha llegado el momento de volver a los dados de nuestro juego.
Si lanzamos dos dados, la apuesta más segura es el 7. ¿Por que?. Podemos considerar el resultado de la tirada como el macroestado del sistema, que es el resultado de una serie de microestados, que definimos como el resultado de cada dado en particular (es decir, el estado termodinámico de cada componente del sistema). Al final, el estado global del sistema es el resultado de los estados particulares de cada uno de sus componentes. Por ello, en la primera ecuación hablábamos que la energía del sistema (macroestado) dependía de N, el número de átomos, cada uno de ellos en un ‘microestado’ determinado.
El número 7 es el que resulta del mayor número de combinaciones de los dos dados, por ello es el que más frecuencia presenta en la distribución.
Cuantos más microestados posibles tenga un macroestado, más probabilidad tenemos de tener el macroestado resultante. Esto se llama probabilidad termodinamica. En el caso del número 7, es el macroestado resultante de seis microestados posibles. Y, lo que nos descubrió Boltzmann es que la mayor entropía tiene lugar en el macroestado al cual conducen el mayor número posible de microestados.
Ecuación de Boltzmann, que, en realidad, fue formulada por Planck.
La ecuación de Boltzmann dice que la entropía es igual al producto de una constante (la constante de Boltzmann, determinada por Planck) por el logaritmo de la probabilidad de ese estado. El principio de Boltzmann nos dice entonces que el aumento de entropía en un proceso es la consecuencia de la transición del sistema del estado menos probable al más probable. En nuestro juego de dados, el estado más probable, el de mayor entropía, es el número 7, y por ello el sistema de dos dados tiene la máxima frecuencia en 7 en su distribución de estados.
Veamos otro ejemplo para ilustrar el principio de Boltzmann:
Si pregunto a los estudiantes, a los que se ha engañado con la idea intuitiva de que la entropía mide el grado de desorden, la mayoría responden que la imagen de la derecha se corresponde con el estado de mayor entropía. Pero NO. La probabilidad de que en una distribución de puntos en la cuadrícula (es decir los microestados) no haya ningún punto al lado de otro es MÍNIMA, y, de hecho, esta distribución no se ha generado al azar. Si distrubimos puntos al azar, siempre va a haber alguno pegado a otro.
El estado de la imagen de la izquierda es, por tanto, más probable, no importa cómo estén ordenados los puntos. Y es más probable, porque por azar resultan mas combinaciones de puntos (microestados) con el mismo número de contactos de unos puntos con otros. Por tanto, la imagen de la izquierda sería el estado de mayor entropía.
Entropía: una medida de la ‘utilidad’ de la energía y la burocracia como análogo
En todo proceso, hay un intercambio energético. Esta energía se invierte en hacer ‘algo’, como mover un motor, o hacer una transformación química. Esta energía que invertimos e intercambiamos la llamamos energía libre.
Pero claro, para que se produzca el proceso, hay que pagar una especie de impuesto: No toda la energía se invierte en hacer algo útil (como mover el motor o transformar moléculas), sino que una parte se «disipa» sin que haga nada de utilidad. Es el término entrópico de la energía libre.
Esta idea de ‘utilidad’ de la energía la podemos ilustrar con una analogía con la burocracia.
Todos odiamos la burocracia. En mi trabajo como profesor, la burocracia requiere una cantidad considerable de esfuerzo y dedicación. Podríamos definir la burocracia como el trabajo que no se invierte en nada productivo, pero se disipa para mantener la estructura. Como veis, volvemos a la idea de entropía como generadora de orden.
La burocracia es entropía que se genera y se disipa. En la disipación de esta entropía reside el mantenimiento de toda una estructura organizativa. Es decir, cuanto más grande y compleja es una organización, más estructura requiere mantener y más burocracia, es decir, disipación de entropía requiere. ¿como aplicamos aquí el principio de mínima producción de entropía de Prigogine?. Muy sencillo: intentamos minimizar el tiempo que dedicamos a la burocracia, maximizando la disipación de ésta. Un sistema biológico funciona de modo parecido, manteniendo así su estructura. El problema es que un sistema disipativo, ya sea un organismo, una estructura o un estado, va a depender de este proceso de generación y disipación de entropía. Ello lleva a que la complejidad vaya aumentando hasta que el sistema no es capaz de disipar más entropía de la que produce, teniendo lugar el envejecimiento y degradación de la organización.
En un estado o empresa es igual: si los requerimientos burocráticos aumentan en exceso, el sistema pierde eficiencia en la disipación de entropía que se acumula, dando lugar a la degradación de la estructura y su muerte. Todos conocemos este proceso cuando estamos sometidos a burocracia paralizante, que hace que sea muy dificil llevar a cabo tareas útiles y todas las estructuras se degradan.
La entropía: una medida de la ‘degradación’ y ‘dispersión’ de la energía
Podemos ilustrar esta idea de degradación de la energía usando una analogía: billetes y moneda fraccionaria
Si tomo un billete de 20 euros y lo cambio en 40 monedas de 50 céntimos, voy a tener el mismo valor monetario (es decir, la energía se conserva). Sin embargo, estoy «esparciendo» o «dispersando» ese valor: he aumentado la entropía (ΔiS>0). Si yo ahora doy una moneda a cada persona que pasa por la calle, ese valor (esa energía) la he «esparcido» y la he expulsado del sistema: he aumentado la entropía del entorno, y he reducido la entropía de mi sistema (ΔeS<0), pero a costa de pagar 20 euros. Cada persona que se ha llevado 50 céntimos, realmente poco puede hacer con ellos. Habría hecho más con los 20 euros, ¿verdad?. Es decir, con el aumento de entropía, al repartir y «esparcir» la energía, ésta se ‘degrada’.
Este proceso, en el que, globalmente, me he quedado igual que antes, pero con 20 euros menos, es un proceso cercano al equilibrio. Ahora imaginad que yo hago lo mismo, pero le pido a cada persona a la que doy una moneda que me suba una cosa que he comprado en el súper y la coloque correctamente en la cocina. Estoy estableciendo unos ciclos y la disipación de los 20 euros lleva a que surja una estructura ordenada en mi sistema. Entonces estaríamos hablando de una estructura disipativa: El empleo de energía y la disipación de entropía, generan la estructura. Imaginad que cada moneda es el CO2 que expulso en cada ‘ciclo’ metabólico, y que el billete original era la glucosa que he consumido en el proceso y tendremos una analogía para relacionar la entropía con el metabolismo. Como con las monedas, el CO2 tiene escaso valor (es una molécula muy estable y con un sobrepotencial muy elevado, que la hace poco reactiva, de ahí la dificultad para convertirla en otros materiales), comparado con la glucosa original, de la que se puede extraer bastante energía. Ahora necesitamos un sistema que recopile las monedas y las convierta de nuevo en billetes de 20 euros. Este sistema son los organismos autótrofos, que, utilizando la luz o la energía química de los minerales, fijan el CO2, convirtiéndolo de nuevo en ‘billetes’ de alto valor: los azúcares, que a su vez se transforman en otros muchos productos.
Esta idea de los ciclos la expresó Harold Morowitz en su Teorema de los Ciclos de Morowitz:
Esto nos permite definir la entropía de un modo intuitivo: es una medida de la dispersión de la energía: cuanto más dispersa, es decir, cuantas más formas tengamos de distribuir la energía entre los componentes del sistema, mayor es la entropía. Lógicamente, si en los productos de la reacción tengo mucho CO2, tengo muchas formas de distribuir la energía entre cada una de las moléculas de CO2; por tanto, tengo más microestados que se manifiestan como el macroestado, y eso aumenta la probabilidad y la entropía.
También decimos que al aumentr la entropía, aumentamos los grados de libertad del sistema. Cuantos más grados de libertad, mayor entropía. Veámoslo con un ejemplo clásico de Lehninger:
¿cual de los tres casos, a, b o c, tiene mayor entropía?. ¿puedes definirlo según el concepto de «estado ordenado» que te contaron? En el caso (a) tenemos dos reactivos separados. Podemos distribuir energía entre las dos moléculas y todas las posibles vibraciones entre sus enlaces. Tened en cuenta que, en la molécula, la energía se distribuye haciendo vibrar y rotar los enlaces, y en el caso (a) hay muchas más formas de hacerlo que en los otros (tenemos más microestados). En el caso (b) tenemos una sóla molécula, pero esta molécula tiene tres enlaces capaces de rotar completamente, con lo que tenemos más grados de libertad, es decir, más formas de distribuir la energía, es decir, más microestados, que en caso (c), en el que tenemos una molécula rígida, por lo que las posibilidades de rotaciones y vibraciones disminuyen. Así, el caso (c) es el de menor entropía y el caso (a) el de máxima entropía. Esto hace que la reacción (a) sea mas lenta que la (c). Mucho más lenta. Y aquí entramos en cómo la entropía gobierna la velocidad de las reacciones y la acción de las enzimas, lo cual es contenido para otro tema.
Intuitivamente, podemos expresar que en el caso (a), podemos ‘esparcir’ la energía mucho mejor que en el caso (c). Entonces, una habitación desordenada no tiene por qué ser el estado más probable, y por tanto, el de mayor entropía. Depende de cómo se distribuya la energía entre sus componentes y del número de microestados que den lugar a ese estado desordenado. Jaque mate, amigos que usais la entropía como excusa para no ordenar la habitación.
Veamos otro ejemplo intuitivo, el de una tostada con mantequilla:
Yico…
Mientras la temperatura se mantenga baja y no proporcionemos calor, estaremos en un estado de baja entropía y el trozo de mantequilla se mantendrá sólido sobre la tostada. Al calentar, ¿qué es lo más probable? que la mantequilla se ‘esparza’ por la tostada. Este será un estado de mayor entropía: las moléculas de la mantequilla se separan (tiene mas ‘libertad’) y distribuyen por la tostada y la energía se reparte, ‘esparciéndose’. Aquí no hay sorpresas: tenemos un proceso irreversible (una vez esparcida la mantequilla, no podemos revertir el proceso), que se mueve, tal como nos decía Boltzmann, hacia su estado de mayor probabilidad. La mantequilla no se ‘desordena’, eso es nuestra percepción de orden y desorden, ya que antes de calentar la mantequilla no estaba formando una estructura cristalina, por ejemplo. Solo era más viscosa, dando la apariencia de ser sólida. Simplemente, la energía se ‘reparte’ siguiendo el camino más probable, aumentando los grados de libertad moleculares, o aumentando el número de microestados. Como la mantequilla es un fluido no compresible, entonces la variación de entropía sólo depende de la temperatura. Al calentarla de A a B, la entropía se incrementa según:
En la ecuación, n es el número de moles y Cp el calor específico. En la tostada siempre estamos cerca del equilibrio termodinámico, por lo que aquí no vamos a tener un fenómeno de estructura disipativa, que resulta de la combinación de reacciones reversibles e irreversibles interconectadas.
Estos conceptos intuitivos que relacionan la entropía no con el orden, sino con la probabilidad y con la distribución de la energía entre componentes del sistema e intercambio con el exterior, son útiles para entender la relación entre entropía, orden biólogico o para entender cómo funcionan las enzimas o la ribozima esencial que da vida a todos los organismos vivos de la Tierra: la peptidil transferasa.
Así, finalmente, la vida, como una estructura ordenada, depende de la generación de entropía y del aumento de entropía del entorno. Consumimos moléculas de alta energía para mantener nuestra estructura y, la entropía del entorno es el precio que hay que pagar por la estructura. Como dijo Schrödinger:
Cuanto más suntuoso es un palacio, mayor es el montón de basura que genera
La emisión de CO2, orina y desechos es esencial para ‘dispersar’ o ‘esparcir’ energía, aumentando la entropía del exterior. Por ello, toda organización basada en el consumo de energía siempre, por definición, es un orden temporal, cambiante y destinado a desaparecer, altamente dependiente de cual es el mecanismo por el que disipa la energía de sus fuentes de energía y del balance entre producción y disipación de entropía. Ya sea una célula, un organismo vivo o una sociedad, todas son estructuras disipativas, temporales y dependientes de la producción de entropía. Debemos estar preparados para ello.
Referencias
Cassani, A., Monteverde, A. & Piumetti, M. (2021) ‘Belousov-Zhabotinsky type reactions: the non-linear behavior of chemical systems’, Journal of Mathematical Chemistry, 59(3), pp. 792–826. doi: 10.1007/s10910-021-01223-9.
De Castro, C. & McShea, D. W. (2022) ‘Applying the Prigogine view of dissipative systems to the major transitions in evolution’, Paleobiology, 48(4), pp. 711–728. doi: 10.1017/pab.2022.7.
Harisch, K. (no date) ‘How the mind comes to life : The physics and metaphysics of dissipative structures’, pp. 1–18.
Hooker, C. (2011) ‘Introduction to Philosophy of Complex Systems: A’, in Philosophy of Complex Systems. Elsevier, pp. 3–90. doi: 10.1016/B978-0-444-52076-0.50001-8.
Howell, L., Osborne, E., Franklin, A. & Hébrard, E. (2021) ‘Pattern recognition of chemical waves: Finding the activation energy of the autocatalytic step in the Belousov-Zhabotinsky reaction’, Journal of Physical Chemistry B, 125(6), pp. 1667–1673. doi: 10.1021/acs.jpcb.0c11079.
Kondepudi, D. K., De Bari, B. & Dixon, J. A. (2020) ‘Dissipative structures, organisms and evolution’, Entropy, 22(11), pp. 1–19. doi: 10.3390/e22111305.
Naderi, M. (2020) ‘On the evidence of thermodynamic self-organization during fatigue: A review’, Entropy, 22(3), pp. 1–32. doi: 10.3390/E22030372.
Ota, K., Mitsushima, S., Matsuzawa, K. & Ishihara, A. (2014) ‘Assessing the environmental impact of hydrogen energy production’, in Advances in Hydrogen Production, Storage and Distribution. Elsevier, pp. 32–42. doi: 10.1533/9780857097736.1.32.
Shanks, N. (2001) ‘Modeling Biological Systems: The Belousov–Zhabotinsky Reaction’, Foundations of Chemistry, 3, pp. 33–53.
Tsuda, I., Yamaguti, Y. & Watanabe, H. (2016) ‘Self-organization with constraints-A mathematical model for functional differentiation’, Entropy, 18(3), pp. 1–13. doi: 10.3390/e18030074.
Científicos e investigadores. Una reflexión contra la excelencia.
escrito por C. Menor-Salvan | 3 octubre, 2023
Alguna vez me han preguntado por qué siempre distingo entre investigador y científico, y cual es la diferencia.
Hay una respuesta corta y rápida: Ser científico es una condición, una forma de vida. El científico lo es por cómo piensa, qué se pregunta y cómo interroga a la realidad. La Ciencia es una forma de gobernar el pensamiento, junto con una gran cantidad de conocimiento. Investigador hace referencia a la aplicación, a la parte práctica. A la acción de investigar. El investigador no tiene por qué ser un científico.
Entonces yo hago la siguiente reflexión, que espero sirva a los nuevos científicos que llegan a mi clase y comienzan en este mundo: Actualmente, a lo largo de la carrera nos educan en la competición y nos inundan con el mito del talento, la excelencia y con la «orientación al logro» (digo actualmente, porque cuando yo estudié no había nada de eso: los profesores, salvo alguna excepción, eran meros autómatas que transmitían una información desde unos papeles amarillos por el uso curso tras curso y con quienes teníamos mínima interacción).
Constantemente estamos sometidos a lo que J. Krishnamurti llamaba la «violencia de la comparación«: el fracaso del otro como medida de mi propio logro. Hay que sacar mejor nota que el otro, hay que conseguir la beca antes que el otro, hay que publicar más artículos que el otro, hay que conseguir el contrato compitiendo con el otro; ser el primero en la clase, la oposición, el primero en publicar algo, etc.
Todos los seres humanos desean poder, riqueza, posición social. El deseo de poder se expresa de muchos modos: está en el profesor, la pareja, en un estudiante respecto de otro. Este deseo de posición dominante es una de las formas de agresividad del hombre. La agresividad y el sometimiento a ella pervierten toda relación a lo largo de la vida. El hombre ha aceptado esto como natural, con todos los conflictos y desdichas que conlleva. Básicamente, en ello se encuentra involucrada la medida -el más, el menos, lo mayor y lo menor- que en esencia implican comparación […]. Ello comienza casi al nacer y continúa a lo largo de la vida-este constante medir el poder, prestigio, riqueza. Esto se fomenta en las escuelas y universidades. Todo su sistema de calificar consiste en una evaluación comparativa. Cuando A es comparado con B, que es brillante, agresivo, inteligente, esa comparación misma destruye a A. Esta destrucción toma la forma de la competencia, la imitación y la conformación a los patrones establecidos por B. Ello engendra antagonismo, celos, ansiedad, miedo, y termina por volverse la condición en la que A vive el resto de su vida, siempre midiendo, siempre comparándose. Esta comparación es uno de los muchos aspectos de la violencia.
– Jiddu Krishnamurti. Cartas a las Escuelas.
El concepto de «excelencia» es distinto en el ámbito científico-académico a como entendemos la palabra en el lenguaje normal. Alguien «excelente» mostraba un conjunto de características mas o menos relativas. Cuando decimos «es una excelente persona», es obvio que es algo relativo al resto de personas. Si decimos que alguien es un «excelente fontanero», claramente, en su profesión, muestra un conjunto de capacidades que nos hace preferirle a otros fontaneros. Pero, pensemos en alguien que es una «excelente persona». ¿cómo cuantificamos eso? ¿haríamos un ranking? ¿cual es la persona más excelente que conoces?. ¿Cómo cuantificamos la excelencia de un fontanero? ¿por el número de obras al año? ¿por las buenas referencias? ¿por la relación calidad-precio?. Seguramente sea difícil medir esa excelencia, incluso para algo aparentemente objetivo, que es cómo trabaja un profesional. Al final es un conjunto de factores muchas veces subjetivos o relativos. Si esto es difícil, ¿vamos un paso más allá y, después de implementar una medida de la excelencia, impedimos que los fontaneros que no la alcancen puedan trabajar?
Nos educan no sólo para cuantificar y valorar el «logro» o el «éxito», la «consecución de objetivos y la excelencia» y en «ser el primero» por encima de todo, sino en la aceptación del sacrificio como algo deseable. Vivimos en una especie de culto al sufrimiento. Se asocia el sufrimiento con ese logro y el disfrute con culpabilidad o pérdida de tiempo. Los investigadores aceptan el sacrificio como parte del trabajo y lo imponen a los más jóvenes. «tienes que ir al extranjero (no por que desees aprender allí, sino como imposición de carrera o para ser mejor que el otro)», «es muy difícil ser investigador y tener familia» «tienes que sacrificar relaciones personales» «tienes que dedicar todo tu tiempo a tu trabajo sin descanso» «si eres investigador, prepárate para pasar penurias económicas y precariedad» «no tengas hijos hasta los 45» «como yo lo pasé mal/trabajé gratis/tuve que exiliarme, tu también debes pasarlo mal/etc.». Hemos llegado a la aberrante asociación «ser investigador» = «sufrir» que aceptamos como algo normal, como parte de la profesión.
Como quien va al gimnasio y asocia el dolor, las agujetas y el cansancio con el avance en sus objetivos. «Cómo me he machacado» dicen. En todo momento se usa una terminología bélica: «hay que luchar por…». Muchas veces pienso en éste perverso pensamiento y recuerdo a mi maestro de Aikido, que no toleraba en el Dojo ninguna expresión de agresividad, dominio, superioridad y ego. Disciplinas como el Aikido o el Yoga nos muestran que la asociación tóxica de dolor y sufrimiento con logro debe terminar: a través de ellas fortalecemos el cuerpo y la mente, mantenemos un cuerpo saludable y mejoramos en el estudio, sin necesidad de competir, ni de sufrir, y llevando una práctica vibrante y agradable con, esencialmente, buenas sensaciones.
En el momento en que te intereses por cuales son «buenos» y cuales «malos» entre tus compañeros, estas abriendo una apertura en tu corazón por donde entrará la malicia. Poner a prueba, competir y criticar a otros te debilita y finalmente te derrota
O-Sensei Morihei Ueshiba
Nos intentan hacer creer que vivimos en una confrontación constante con nuestros compañeros y con nosotros mismos, como seres imperfectos en constante test y prueba, intentando mejorarse y ser mejores que otros. Sea lo que sea que signifique eso. Nos presionan para ser «primero», para ser «antes que», en definitiva, para valorar nuestro éxito en la medida del fracaso que perciben los que están en la lista o el ranking tras nosotros. Durante todo el tiempo nos lavan el cerebro con términos como «resiliencia», «talento» y «excelencia». Estas tres palabras, indicadores claros de que estamos ante un idiota postmoderno, deberían hacernos saltar las alarmas: «resiliencia» es la capacidad para recuperarse de una agresión o perturbación. ¿debemos prepararnos entonces para ser maltratados?. Si alguien os pide «resiliencia» como cualidad para un trabajo, tened por seguro que os van a tratar mal.
Pero vamos a revelar una parte importante del engaño: «talento» y «excelencia» son los términos usados por burócratas y políticos para justificar la explotación, la competición, la falta de oportunidades y financiación y la falta de derechos. «Sólo concedemos X contratos o proyectos, porque estamos fomentando la excelencia y el talento», cuando en realidad quieren decir «Sólo concedemos X contratos porque no queremos invertir más en Ciencia, os haremos creer que nos basamos en la excelencia, sea lo que sea eso, pero solo vamos a financiar lo que queremos/creemos conveniente/a quien hemos financiado ya previamente y ha tenido resultados (el efecto Mateo)». Cuando en una convocatoria de proyectos o contratos incluyen la palabra «excelencia», sólo os están diciendo que hay un pedazo de pastel muy pequeño y esperan que los comensales peleen como perros rabiosos por él. Son un engaño, como toda la cultura del talento. Palabrería que crea un ambiente tóxico, lleva a la infelicidad, la frustración y la decepción. Y la frustración o la aceptación del propio fracaso lleva a la desmotivación y a acumular más fracaso según los parámetros de quienes nos están aplicando sus medidas de excelencia. En el momento en el que aceptamos el fracaso, dejamos de esforzarnos. «¿para qué?» es una pregunta habitual, «si no voy a conseguir esa plaza o proyecto».
Estos sentimientos negativos abundan en los jóvenes científicos y son contrarios a los que deben tenerse al hacer Ciencia: el ambiente en un laboratorio debe ser relajado y motivador. La exploración de la Naturaleza es y debe ser gozosa, no debe ser una fuente de sufrimiento o estrés. Un científico no está interesado en competir, en lograr, en el «talento». Hacemos ciencia porque con ella aprendemos cosas, tenemos experiencias interesantes, vemos la Naturaleza bajo otros ojos y, en definitiva, tenemos una vivencia. No importa lo inteligente que seas, ni tu talento, sea lo que sea eso. La Ciencia es una forma de manejar nuestro pensamiento y enfrentarnos a la realidad. La Ciencia es un modo de relacionarnos con el mundo. Es exploración, aprender y enseñar. Descubrir y transmitir. Parafraseando a Karl Popper,la Ciencia nos interesa porque queremos saber algo del enigma del mundo en que vivimos y del otro enigma del conocimiento humano de este mundo.
Un científico intenta dominar el arte de aprender y, los que tienen la suerte de ser al mismo tiempo científicos y profesores de ciencias, tienen la doble tarea de practicar el arte de aprender y el arte de enseñar.
Pero daré a conocer lo poco que he aprendido para que alguien mejor que yo pueda atisbar la verdad y, en su obra, pueda probar y criticar mi error. Así, me regocijaré a pesar de todo de haber sido un medio a través del cual salga a la luz la verdad.
Alberto Durero
Un investigador os hablará de cuantos «papers» publica y sus cuartiles, cuantos fondos y proyectos consigue, de sus logros, de su talento, de líderes y liderar (otra palabreja postmoderna de moda), de la cantidad de charlas que da y de los sitios y colegas prestigiosos con los que se codea. Vivirá observando baremos, índices de impacto, métricas y rankings. Un investigador no desea el error, porque, durante toda nuestra enseñanza, se nos enseña a temer y odiar el error. A ridiculizar o castigar a quien se equivoca. Un investigador buscará destacar sobre el otro, y aprovechará cualquier posibilidad de promocionarse. Es a lo que nos han enseñado y forma parte del trabajo del investigador científico actual.
Durante la celebración del sexagesimo aniversario de Max Planck, en 1918, Einstein dijo que en el templo de la ciencia hay tres tipos de personas. Muchas se dedican a la ciencia en razón del goce de su poder intelectual superior; para ellos, la investigación es una especie de deporte que satisface su ambición personal. Una segunda clase de investigadores se dedica a la ciencia para conseguir fines exclusivamente utilitarios. Pero, en lo que respecta a la tercera: si el ángel del Señor viniera y sacara del templo a todas las personas que pertenecen a esas dos categorías, quedarían unas pocas personas, entre ellas Planck, y ésta es la razón por la que lo queremos.
Edward O. Wilson. Consiliencia o la unidad del conocimiento
Un científico mas bien os hablará de las cosas que ha visto y aprendido; cuando habla de publicaciones, explica lo que ha publicado, no de dónde lo ha hecho o cuanto ha publicado, o de su impacto; habla de lo que ha experimentado, lo que observa, sus ideas, lo que le ha fascinado en su exploración de la vida y la naturaleza. No le interesará dónde has publicado o con quién trabajas, o si has estado en un centro extranjero de nombre sonoro, sino qué te apasiona, qué preguntas e ideas tienes y qué opinas. Un científico abraza el error, porque el error, que puede saber amargo, madura en el delicioso fruto de la Ciencia y el aprendizaje.
La Ciencia se ha construido sobre la refutación de ideas previas. En el momento en el que el investigador trabaja en la Ciencia en un ambiente de violencia y confrontación, como ocurre en cualquier batalla, un error puede ser fatal. Así, es fácil ver que la construcción y el trabajo de la Ciencia no es compatible con los sentimientos de estrés, competitividad, frustración y miedo al fracaso a los que se someten a los científicos. En el momento en que eres científico (no investigador), no existe el fracaso; existen, mas bien, resultados negativos, refutación de tus ideas previas, resultados que no puedes interpretar y limitaciones, falacias y paradojas cognitivas. Todos estos factores son tan importantes o más para la Ciencia que la verificación de una conjetura o teoría. Es importante por ello, que el científico deje de lado el orgullo y acepte con humildad su verdadera posición de observador y transmisor de conocimiento. Naturalmente, «científico» e «investigador» no son dos categorías absolutas y separadas. Podríamos decir, si acaso, que «investigador» es el trabajo y «científico» una condición, una forma de vida. Todos navegamos entre las dos aguas, salvo excepciones (investigadores que son meros trabajadores y tienen poco de científico y, mas raramente, científicos idealistas empeñados en alejarse de la realidad). A mi me parece, a veces, complicado encontrar el equilibrio.
Quienes os digan que hay que competir, que todo esto que digo es incompatible con tener objetivos, con el esfuerzo, con tener resultados y eso que llaman «éxito», sólo os están engañando o manipulando. Se puede ser un científico y tener disciplina, marcarse objetivos o realizar un gran esfuerzo. La diferencia es que éste esfuerzo no estará motivado por la competitividad, por el miedo al fracaso o por la búsqueda del logro, sea cual se este (ya sea salvar a la humanidad, publicar con alto índice de impacto o satisfacer el placer de recibir elogios), sino motivado por la búsqueda del aprendizaje, por comprender un poquito más un pequeño aspecto de la realidad, por responder y/o plantear preguntas; y también, por qué no, realiza un gran esfuerzo por el pundonor del trabajo bien hecho, por el arte, digamos. Para el científico no hay ciencia pequeña o irrelevante. El investigador desea el impacto, porque le aportará fama y prestigio. Para el científico, toda la ciencia es necesaria e importante. Un dato que aportas puede que no cambie ningún paradigma, pero dentro de 5, 10 o 20 años puede ser el factor clave de un gran descubrimiento. Hay muchos ejemplos de ello, y los profesores deberían hacer hincapié en esa «ciencia pequeña» tan importante. La calidad de la ciencia no se mide con su impacto o según dónde se publica, sino según el modo en que ha sido realizada, según su técnica y discusión y según la veracidad y honestidad de su resultado, da igual si es un «pequeño resultado» o no. Es la velocidad y el simplismo de la sociedad moderna lo que lleva a juzgar la calidad de un resultado científico por el título de la revista en que se publica o según cuanto se cite en los medios.
Eso se aplica a cualquier Ciencia: saber un poco más del mundo en que hemos vivido y el universo del que formamos parte.
Según todos estos factores, un investigador quizá pueda ser mejor que otro. No lo se. Quizá podamos inventar métricas y rankings para decidir artificialmente quien es absolutamente mejor o peor investigador en base a una métrica relativa. Igual que las notas de clase: ¿será mejor biólogo molecular quien haya sacado matrícula en la asignatura de biología molecular?. Igual, simplemente, en esa métrica particular obtuvo un valor mas alto, pero, ¿es mejor o peor? ¿en qué y en base a qué? Igual fue pura casualidad. Igual es un desastre en el laboratorio. O igual no. Tratar de decidir quién es mejor o peor científico es tan ridículo como observar un parque en el que juegan grupos de niños y tratar de hacer un ranking de los que mejor juegan. «Juanito ha subido al tobogán un 20% más que Carlitos». «Anita sube más arriba en el columpio que el 75% restante de los niños». ¿no os suena ridículo?.
Determina cual de estos niños juega «mejor» y luego establece un ranking de «excelencia». A quien esté el último, no le dejes jugar en el parque. ¿podríamos hacer algo así? ¿cómo?. Con la Ciencia ocurre algo similar.
Podemos intentarlo, claro, pero ellos estarán ajenos a ello, y, si intentamos que entren en nuestro juego de rankings (dando de merendar o un juguete, o permitiendo jugar en el parque sólo al primer cuartil en el número de veces que suban a lo más alto de determinado columpio), solo lograremos arruinar su tiempo de juegos, desmoralizarles y convertir el parque en un campo de batalla. He visto, a lo largo de mi carrera, personas moralmente deshechas por su trabajo como investigadores. Personas inteligentes y con muchas ideas y pasión abandonando la Ciencia. He visto agresiones e intentos de asesinato. Suicidios. Yo mismo he sufrido estrés y depresión causada por mi trabajo. Esto es lo que provocan aquellos que hablan de «talento», «excelencia», «resiliencia», «competitividad», etc. ¿es esto lo que queremos para nosotros, nuestros alumnos y compañeros? ¿donde queda, en éste ambiente de «competitividad por la excelencia» la amabilidad y la bondad?
La bondad solo florece en libertad. No puede hacerlo en el suelo de la coacción bajo ninguna de sus formas, ni bajo compulsión, ni es el resultado de la búsqueda de recompensas. No puede existir cuando hay temor.
Jiddu Krishnamurti
Alguien podría decirnos, «es el sistema». Que no os mientan: el sistema lo hacen personas como vosotros y yo. En nuestra mano está cómo queremos que sea. Podéis enfocar vuestra carrera de dos formas: como un investigador que quiere ser el primero o como un científico que quiere descubrir. En ambos casos haréis básicamente lo mismo (mucho trabajo, burocracia, participar en concursos por plazas y proyectos, intentar publicar papers lo mejor posible…etc.) La diferencia es que, en el primer caso, es muy probable que viváis una vida de frustración, de estar pendientes de rankings y baremos, de lo que hacen otros, de alimentar el ego, de sufrir y hacer infelices a otros. En el segundo caso es posible que lo paséis mal a veces, que sea difícil, que os disgustéis. Pero siempre encontraréis gozo, motivaciones y alegrías. En el primer caso viviréis pensando en el fracaso, en el segundo caso no hay fracaso ni éxito, simplemente viviréis una vida gozosa como científicos, independientemente de dónde estéis en el ranking. Y, recordad algo muy importante: en el mundo de la Ciencia casi todo el mundo es inteligente, y «excelente». Si queréis marcar la diferencia, sed amables, humildes y generosos. Sed científicos, no investigadores.
Los actores se ponen maquillaje y representan la belleza y la fealdad; pero cuando se termina la obra, ¿Dónde están la belleza y fealdad?. Los jugadores de ajedrez compiten por ser los primeros e intentan aventajarse unos a otros en sus movimientos; pero cuando se acaba el juego y se retiran las piezas, ¿Dónde queda entonces la competición?
¿Por qué aumenta la infectividad del coronavirus?: Un sencillo experimento virtual con la ‘variante británica’ del SARS-CoV-2
escrito por C. Menor-Salvan | 3 octubre, 2023
por C. Menor-Salván
En los medios y redes sociales ha causado una gran alarma la aparición, reportada en diciembre de 2020 en Gran Bretaña, de una nueva variante del virus SARS-CoV-2. Esta variante, denominada B.1.1.7, contiene varias mutaciones puntuales respecto de la variante original que se extendió en febrero-marzo de 2020. Según los datos clínicos disponibles, la variante B.1.1.7 tiene mayor infectividad y velocidades de transmisión. Desde diciembre, según el COVID-19 UK Genomics Consortium, más del 50% de los nuevos contagios son de ésta variante, que presenta una transmisibilidad incrementada entre un 50 y un 70% respecto de la variante original que llamaremos WT (por ‘wild type’). Como es lógico, la aparición de variantes con mayor transmisibilidad crean alarma, pero son perfectamente esperables, dado que estamos forzando el proceso de evolución viral: mediante confinamientos y medidas anticontagio, creamos una selección artificial de las variantes más infectivas, que prevalecen sobre las variantes originales.
La infectividad viral es una consecuencia de varios aspectos moleculares del virus; uno de ellos es la afinidad por el receptor celular, que una proteína del virus reconoce, y al que se une, iniciando el proceso de infección. La proteína viral que reconoce el receptor en la célula que va a infectar es la proteína de la espícula. Como si de un puzzle molecular se tratara, la espícula viral ‘encaja’ en el receptor. Os remito a la Noticia Nº 59 para una introducción general al SARS-CoV-2
La proteína de la espícula, que contiene un dominio o región, que interacciona con el receptor de la célula.
Como es lógico, si modificamos la proteína de la espícula de modo que mejore la estabilidad de su interacción con el receptor, aumentará la infectividad. ¿como?. Para entenderlo, hay que tener en cuenta que un virus es un agregado supramolecular. Digamos que es una gran molécula heterogénea. No es un ser vivo. Si el virus tiene mayor afinidad por su receptor, quiere decir que la infección se producirá a una concentración menor. Dicho de otra manera, si aumentamos la afinidad del virus modificado por el receptor, se producirá una infección con una cantidad de virus menor que los necesarios con el virus sin modificar. Y, posiblemente, este proceso está detras del efecto de la variante UK B.1.1.7: una de sus mutaciones se encuentra precisamente en la región de la espícula que interacciona con el receptor de la célula a infectar.
En ésta entrada voy a mostraros como nosotros mismos podemos ver éste efecto: voy a tomar los datos de la estructura PDB 60MJ y, a partir de ellos, voy a:
Calcular las interacciones por puentes de hidrógeno entre la espícula y el receptor. Estas interacciones determinan la estabilidad el complejo espícula-receptor, y, como es lógico, cuando más puentes y más fuertes, mayor estabilidad = mayor afinidad =mayor infectividad. También observaremos otras interacciones evidentes, como el stacking pi: un tipo de enlace que se produce entre aminoácidos aromáticos.
Voy a introducir la mutación N501Y de la ‘variante británica’, que implica el cambio de la asparagina 501 de la espícula por una tirosina, justamente en la región de interacción entre la espícula y el receptor.
Después, usando el software, el ordenador calculará la estructura más estable resultante de ése cambio y veremos si la mutación introduce la aparición de nuevas interacciones. Si la mutación implica la aparición de nuevos enlaces, esto explica fácilmente el aumento de infectividad de la ‘variante británica’.
Este procedimiento lo voy a llevar a cabo usando los softwares UCSF Chimera, Pymol y Chem3D.
La mutación N501Y de la ‘variante británica’ del SARS-CoV-2 introduce nuevas interacciones y podría aumentar la afinidad por el receptor y la infectividad
Como hemos comentado, la unión entre la espícula viral y el receptor desencadena el proceso infectivo. Esta unión se lleva cabo gracias a que se establecen una serie de interacciones no covalentes entre las dos proteínas: puentes de hidrógeno e interacciones tipo apilamiento entre anillos aromáticos. Cuanto más fuerte sea la unión, mayor estabilidad tendrá el complejo y, por tanto, mayor afinidad tendrá el virus por el receptor. Al aumentar la afinidad, aumenta la infectividad del virus, dado que se requiere una inoculación de menor cantidad de virus para producir una infección efectiva. Si, por ejemplo, el virus WT necesita (número arbitrario) introducir 1000 unidades de virus en el hospedador para producir una infección, al aumentar la afinidad por el receptor ese número se puede reducir a 500-300.
Interacción entre la glicoproteína S o espícula viral y la proteína receptora ACE2 de la célula, que a su vez forma un complejo con el transportador transmembrana de aminoácidos B0AT1. En las siguientes figuras, nos centraremos en la zona recuadrada, donde se produce la interacción espícula-receptor. Imagen tomada de Rynkiewicz et al. (2021)
Centrándonos en las estructuras del dominio RBD de la espícula y de la proteína ACE2 (recuadrados en la figura anterior), vamos a ver el resultado que obtenemos usando los datos de la variante original del virus:
Vemos la estructura molecular del dominio RBD, en azul. Este dominio es la parte de la espícula que se une al receptor celular, la proteína ACE2. Las líneas azules representan los puentes de hidrógeno entre aminoácidos. Aquí tienen especial protagonismo el Gln 493 y la lisina 417, que forman puentes con el receptor. La fenilalanina 486 da lugar a un apilamiento pi con una tirosina del receptor. Esta interacción no existía en el SARS de 2003, y es una de las que explica la mayor afinidad (e infectividad) del SARS-CoV-2. En amarillo veis la Asn (asparagina) 501, que ya en su momento se señalaba como un aminoácido potencialmente interesante en la estructura. Vamos a ver qué ocurre cuando introducimos la mutación:
Fe de errata: Donde dice «E50Y» debe decir «N501Y»
La introducción de la mutación, con la nueva tirosina en la posición 501, tiene consecuencias que son sutiles pero de gran importancia. Primero, aparecen nuevas interacciones entre el nuevo aminoácido y el receptor: nuevos puentes de hidrógeno y un enlace por stacking pi entre la tirosina 501 y una tirosina del receptor. Por otro lado, interacciones que ya existían previamente disminuyen su distancia. Los puentes de hidrógeno entre las lisina 417 y la glutamina 493 se hacen más cortos (y por tanto mas fuertes, ya que es una interacción electrostática). Como resultado la proteína mutante se une con mayor estabilidad al receptor, ya que aumentan el número de puntos de atracción electrostática y disminuye la distancia de las que ya existían. Esto da lugar a una unión más estable, y, por tanto, a un aumento de la afinidad. Esto se traduce en la práctica en un aumento de la infectividad de la variante.
Las representaciones de proteínas son algo abstractas. Para que el lector pueda ver más intuitivamente cómo se produce la unión, vamos a ver las superficies de las moléculas de proteína. Estas interaccionan ‘encajando’ y uniéndose por las interacciones electrostáticas, de modo similar a como si uniéramos unas piezas con imanes.
Interacción entre el dominio de unión de la proteína de la espícula y el receptor. El receptor es una enzima del tipo zinc- metaloproteasa, y puede observarse el gran surco del centro activo, donde encaja la angiotensina.
En cierto modo, el virus y su receptor interaccionan de modo similar a las piezas de un conector magnético, pero con una interacción electrostática en lugar de magnética.
La fuerza y geometría de las interacciones electrostáticas determina la estabilidad de la unión entre el virus y su receptor, y, por tanto, su afinidad e infectividad. La acción de los anticuerpos se basa en un principio similar. Una pregunta habitual es si ésta nueva variante supondrá una pérdida de eficacia de las vacunas o mejorará la evasión inmune del virus. Todavía queda mucho trabajo para los científicos, pero los primeros datos indican que ésta variante no alterará la eficacia de la vacuna ni la respuesta inmune, al ser mutaciones puntuales que no afectan al reconocimiento de los anticuerpos.
Por otro lado, las modificaciones en el dominio de reconocimiento de receptor en la espícula viral, tampoco implican un aumento de la agresividad del virus, por lo que el aumento de infectividad no va asociado a un aumento de gravedad de las infecciones. Como digo, aún queda mucho que investigar y que aprender y todo puede ir cambiando, pues la pandemia evoluciona más rápido que la capacidad de los científicos para obtener resultados y avances.
Estos sencillos resultados que he mostrado, obtenidos mediante un no menos sencillo análisis computacional de datos de estructuras de proteínas, no son mas que una aproximación muy sencilla. Un estudio mas complejo y riguroso requeriría muchos mas medios y tiempo de los que dispongo, pero espero que sirva para entender la mecánica molecular que hay tras la infección del coronavirus.
Referencias
Kupferschmidt, K. Fast-spreading UK virus variant raises alarms. Science (New York, NY), 371(6524), 9-10.
Lauring, A. S., & Hodcroft, E. B. Genetic Variants of SARS-CoV-2—What Do They Mean?. JAMA.
Rynkiewicz, P., Babbitt, G. A., Cui, F., Hudson, A. O., & Lynch, M. L. A comparative survey of Betacoronavirus binding dynamics relevant to the functional evolution of the highly transmissible SARS-CoV-2 variant N501Y. bioRxiv, 2021: https://doi.org/10.1101/2020.09.11.293258
Tang, J. W., Tambyah, P. A., & Hui, D. S. (2020). Emergence of a new SARS-CoV-2 variant in the UK. Journal of Infection.
Volz, E., Mishra, S., Chand, M., Barrett, J. C., Johnson, R., Geidelberg, L., … & Ferguson, N. M. (2021). Transmission of SARS-CoV-2 Lineage B. 1.1. 7 in England: Insights from linking epidemiological and genetic data. medRxiv, 2020-12.
Venenos, el arma bioquímica: El caso de la avispa esmeralda
escrito por C. Menor-Salvan | 3 octubre, 2023
La avispa usa la Bioquímica para atrapar a su presa sin violencia ni dolor
No hay víspera del día de difuntos sin cuento de miedo. Y no hacen falta fantasmas ni fenómenos paranormales para ello, ya que la Naturaleza nos provee de historias que podrían inspirar muchas películas truculentas. Es el caso de la relación mortal entre la avispa esmeralda (Ampulex compressa) y las cucarachas.
La avispa parasitoide
Las avispas-joya o avispas esmeralda (género Ampulex) son un caso único desde el punto de vista del comportamiento y de la Bioquímica. Viven en una amplia distribución, fundamentalmente en África, Sur y Sudeste asiático, América e islas del Pacífico; aunque en Europa también se encuentran, son poco abundantes.
Estas avispas son parasitoides, insectos que viven su vida adulta independientemente, pero que durante su estado larval parasitan y destruyen otro insecto hospedador. Normalmente, el adulto captura, ataca o inmoviliza al hospedador, sobre el que deposita un huevo. El hospedador servirá de refugio y alimento a la larva hasta su transformación en adulto, implicando la muerte del hospedador, normalmente lenta y macabra. Un ejemplo de comportamiento parasitoide lo tenemos en el cine: las películas Alien (Ridley Scott, 1979) y Aliens (James Cameron, 1986).
La avispa esmeralda se aproxima cuidadosamente a su víctima, una cucaracha de tamaño y fuerza muy superior a ella. Cuando localiza la posición de ataque adecuada, la avispa se lanza sobre la víctima, clavando su aguijón justo en una zona específica de su tórax, paralizando un par de las patas de la cucaracha. Esta parálisis dura un par de minutos, suficiente para la avispa, que se agarra fuertemente a lo que podríamos llamar el cuello de la víctima e inyecta veneno de nuevo, esta vez con gran precisión, directamente en el cerebro de la cucaracha. La cucaracha entonces entra en un estado placentero, que dura unos 20 minutos, similar a un post-coito o el que se produce tras una comida abundante, en el que se dedica a limpiarse cuidadosamente, mientras la avispa espera. Tras ese tiempo, el veneno ha ido haciendo su efecto completo y la cucaracha pierde toda voluntad para realizar cualquier movimiento, reflejo de huida o defensa. Entonces, la avispa se acerca y mordisquea las antenas de la víctima (se cree que es un test para comprobar que el veneno inyectado haya hecho efecto, pero no sea tanto como para matar a la cucaracha). La víctima, a partir de ese momento, solo se moverá si la avispa, a la que ha dejado de ver como una amenaza, tira de ella. Dócilmente y en un estado cerebral inducido por unos niveles muy altos de dopamina (similar a los que tienen los enamorados o los individuos muy motivados), la cucaracha irá donde la avispa la lleve. Una vez en la guarida de la avispa, ésta coloca un huevo justo debajo de una de las patas y en una zona con acceso a tejidos blandos.
Avispas esmeraldas picando su aguijón en el cerebro de cucarachas (arriba) y avispa conduciendo a su víctima al nido, donde colocará el huevo del que saldrá la larva que consumirá los tejidos de la cucaracha aún viva (imágenes del naturalista italiano Emanuele Biggi)
En unos tres días, la larva eclosionará del huevo y empezará a succionar la hemolinfa de la cucaracha mientras segrega un cóctel de compuestos antimicrobianos, hasta que, en la siguiente fase, una vez desarrolladas las mandíbulas, entrará dentro de su cuerpo y devorará lentamente sus tejidos internos, de modo selectivo para mantener viva a la cucaracha el mayor tiempo posible en una tortura que puede durar 8 días. Tras ese tiempo, tiene lugar la fase de pupación, y en cuatro semanas una nueva avispa adulta saldrá de los restos secos de la cucaracha. Es interesante que el efecto del veneno en el cerebro de la cucaracha dura unos 8 días (suficiente para cubrir el proceso). En experimentos en los que se retira el huevo de una cucaracha atacada, el animal se recupera y vuelve a la normalidad en ese periodo.
Avispa adulta emergiendo de los restos de una cucaracha muerta. Foto: Emanuele Biggi.
El veneno: un arma bioquímica perfecta
Este mismo año (2018), gracias a dos extraordinarios trabajos científicos se ha encontrado y caracterizado uno de los componentes únicos y esenciales del veneno de la avispa esmeralda: el péptido ampulexina. Además de estos nuevos compuestos, el veneno de la avispa contiene otra sustancia característica, que es la dopamina, neurotransmisor que, entre otras funciones, activa los circuitos cerebrales de placer y recompensa y tiene un efecto excitante que ayudará a mantener a la cucaracha con vida. El cóctel de ampulexinas y dopamina, inyectado directamente en el cerebro de la cucaracha, anula totalmente su voluntad, convirtiéndola en un dócil, amigable y enamorado sirviente de la avispa, que la seguirá ciegamente hacia un destino mortal: ser alimento de su carnívora descendencia.
Ampulexina 1 (izquierda) y ampulexina 2 (derecha), un pequeño péptido que forma una alfa-hélice y es el componente esencial y único del veneno de la avispa esmeralda. La ampulexina provoca hipocinesia (pérdida de la capacidad de moverse a voluntad) y hace que la cucaracha sólo se mueva cuando la avispa la dirige. Imagen tomada de Moore et al., Biochemistry, 2018, 57 (12), pp 1907–1916 DOI: 10.1021/acs.biochem.7b00916
El análisis mediante proteómica ha revelado que el veneno de la avispa contiene la ampulexina como uno de los componentes esenciales y únicos, característico de esta avispa. Además contiene otros péptidos no identificados (posiblemente relacionados en función, pero que pueden dar sorpresas) y otros componentes, comunes en venenos de insectos, arañas, escorpiones y serpientes. Los venenos de éstos animales, a pesar de su distancia evolutiva, tienen sorprendentes características comunes.
Proteínas del veneno de la avispa esmeralda. Imagen tomada de Arvidson, R., Kaiser, M., Lee, S. S., Urenda, J. P., Dail, C. J., Mohammed, H., … & Adams, M. E. (2018). Parasitoid Jewel Wasp Mounts Multi-Pronged Neurochemical Attack to Hijack a Host Brain. Molecular & Cellular Proteomics, mcp-RA118.
Un componente habitual, muy abundante en el veneno, es la hialuronidasa. Esta enzima, componente importante en los venenos de abejas, avispas, arañas, serpientes y otros animales venenosos, tiene como función destruir el ácido hialurónico, degradando la matriz extracelular de los tejidos. En la abeja esmeralda, la hialuronidasa facilita la penetración en el cerebro de la ampulexina, haciendo que ésta tenga efecto. La hialuronidasa, inyectada en gran cantidad en el veneno de arácnidos, contribuye a licuar los tejidos de la víctima, que luego son succionados por la araña. Las células cancerosas humanas también liberan hialuronidasa, que facilita la invasión de los tejidos por los tumores malignos. Es curioso que la hialuronidasa humana es extremadamente similar a la hialuronidasa del veneno de abejas:
Superposición de las estructuras PDB 1FCV y PDB 2PE4 (hialuronidasa de abeja y de tumores humanos) mostrando la gran similitud de las estructuras. Las líneas gruesas coloreadas indican las partes homólogas de la proteína. El análisis estructural a partir del Protein Data Bank indica que son similares con alta significatividad e idénticas en un 30% de la secuencia. (Imagen C. Menor-Salván-Protein Data Bank)
La similitud estructural y de secuencia de la hialuronidasa de los humanos y el veneno de abejas y avispas es sorprendente e indica que provienen del mismo ancestro común. En cierto modo, los humanos conservamos la «programación genética» para generar un veneno similar al de insectos, arañas y serpientes.
Otro componente importante del veneno de nuestra protagonista (y de todos los insectos que pican) es la PLA2 o fosfolipasa A2. Esta enzima destruye los fosfolípidos y uno de sus efectos es provocar una respuesta inflamatoria y dolorosa, al liberarse ácido araquidónico de las membranas celulares atacadas por el veneno. La PLA2 es una de las responsables del dolor asociado a la picadura de una avispa. Curiosamente, las abejas y avispas tienen una pequeña proteína llamada melitina, que acentúa la acción de la PLA2 y provoca un dolor intenso; esta proteína, al parecer, no se ha encontrado en el veneno de la avispa esmeralda (al menos no figura en los análisis más recientes). Es posible que esto contribuya a que la picadura sea menos dolorosa, lo cual ayudaría a que su víctima no se revuelva contra ella y a que el veneno ejerza su efecto hipnótico.
Esto que ha mostrado tan sólo es la punta del iceberg. En realidad, la composición de los venenos de insectos es muy compleja, contiene cientos de sustancias, muchas de ellas desconocidas, de modo que la acción del veneno es el resultado de todos ellos juntos. Muchas de éstas sustancias desconocidas podrían ser, algún dia, parte de tratamientos farmacológicos, lo que nos lleva a preguntarnos:
¿podría tener alguna utilidad éste veneno?
Alguien podría preguntarse: ¿esta mezcla de ampulexina y dopamina podría utilizarse para «zombificar» a los humanos, anular su voluntad y llevarlos donde uno quiera?.
Es cierto que algunos componentes del veneno de avispa y abeja tienen potencial farmacológico. Por ejemplo el mastoparán, péptido que se encuentra en el veneno de avispas comunes, puede tener uso terapéutico como antiviral, antitumoral o en el tratamiento de la enfermedad de Chagas, gracias a su efecto tripanocida. La melitina, que provoca el dolor de la picadura de avispas y abejas, tiene efectos antitumorales muy prometedores. Poco a poco se van viendo más péptidos derivados del veneno de las avispas con actividad farmacológica, un campo de investigación reciente y que aún tiene que superar bastantes barreras.
Las temidas avispas, que no suelen despertar mucha simpatía, podrían ser una valiosa fuente de nuevos fármacos. Los venenos de las avispas ya tienen aplicaciones terapéuticas.
En el caso de las nuevas ampulexinas, es difícil que alguien pudiera usarlas de modo maligno para anular la voluntad en humanos. Hay que tener en cuenta que la avispa esmeralda inyecta el veneno directamente en el cerebro de la víctima, en una maniobra de precisión. La anatomía y fisiología del cerebro humano posiblemente no permita que las ampulexinas actúen de esa forma y, si lo hicieran, para conseguir convertir a un humano en zombi, el cóctel ampulexina-dopamina y todas las proteínas necesarias para que funcione bien, no puede ser inyectado o ingerido, sino que debería ser liberado en una zona específica del cerebro con precisión. De momento, la televisión, la política y la religión consiguen un efecto similar en humanos al que produce la avispa esmeralda en su víctima, y sin necesidad de complejos cócteles bioquímicos.
Robin Williams y el plegamiento de proteínas.
escrito por C. Menor-Salvan | 3 octubre, 2023
El suicidio del actor Robin Williams en 2014 conmocionó al mundo del cine y a todos los que nos gustaba como actor y disfrutamos muchas de sus películas. A mi me gustó especialmente en El Club de los Poetas Muertos, El Indomable Will Hunting y Despertares, donde dió vida al neurólogo, gran amante de la Química y divulgador de la Ciencia que fué Oliver Sacks.
Robin Williams, encarnando a Oliver Sacks, hablando sobre la Tabla Periódica a Robert de Niro en Despertares
Algún tiempo después, su familia reveló que Robin Williams padecía un tipo de demencia, asociada a la enfermedad de Parkinson, llamada demencia con cuerpos de Lewy. Según cuentan, esta enfermedad degenerativa pudo estar detras del suicidio de Williams. Es muy complejo hablar sobre la demencia con cuerpos de Lewy en una entrada de blog, por lo que solo voy a dar unas pinceladas, centrándome en un aspecto fundamental: la importancia de la estructura de las proteínas.
Fijate en ésta imagen:
La gran mancha redonda oscura es un cuerpo de Lewy incluido en una célula de una parte del cerebro de un paciente con demencia con cuerpos de Lewy. Estos cuerpos son masas de proteínas agregadas. Estas masas de proteínas forman en primer lugar unas estructuras que se denominan agresomas (de agresión). La formación de éstos cuerpos es una consecuencia de las propiedades de las proteínas y aquí entramos en el meollo de la cuestión: las proteínas, para realizar su función tienen que plegarse dando lugar a unas estructuras tridimensionales características. Estas estructuras tridimensionales son las que dan sentido y funcionamiento a las proteínas. Lo malo es que muchas proteínas tienden, de modo natural, a plegarse «mal», siguiendo su tendencia natural. Al fin y al cabo una proteína es un tipo de poliamida, como el nylon de las fibras de tejidos sintéticos. Y, como esas fibras, las proteínas tienden a formar fibras insolubles y resistentes. Cuando una proteína en la célula se pliega mal, la célula la marca como defectuosa, para eliminarla antes de que pueda provocar daños. Este marcaje se hace con una pequeña proteína llamada ubiquitina. Las proteínas marcadas se dirigen hacia un sistema de eliminación llamado proteasoma.
Una vista simplificada del sistema de eliminación de proteínas defectuosas. Las proteínas mal dobladas pueden formar agregados insolubles, que se almacenan en el IPOD, o bien se marcan (ubiquitina) y se ponen en «cuarentena» para su revisión o eliminación en el JUNQ. Si las proteinas marcadas se agregan, se «tiran al punto limpio», llamado agresoma. Cuando las células tienen defectos para procesar el «punto limpio» agresoma, éste crece y se convierte en un cuerpo de Lewy, produciéndose la enfermedad. Imagen tomada de M Takalo, A Salminen, H Soininen, M Hiltunen, A Haapasalo, Am J Neurodegener Dis 2013;2(1):1-14.
Cuando las proteínas marcadas como defectuosas se agregan, la célula las amontona en una especie de contenedor, o «punto limpio», que es el agresoma, para evitar que provoquen daños en la célula. La célula entonces elimina la basura de éste punto limpio en un proceso llamado autofagocitosis.
Si falla el proceso de limpieza del «punto limpio» por un defecto, la basura se acumula, forma una masa intratable y el agresoma se convierte en cuerpos de Lewy.
Como vengo diciendo, todo este proceso depende de cómo se doblen las proteínas y de la costumbre natural de las proteínas para formar agregados insolubles de fibras (como las poliamidas que se usan en tejidos). En el caso de la enfermedad que padecía Robin Williams, una de las proteínas responsables de la formación de los cuerpos de Lewy y de la demencia es la alfa-sinucleína. En condiciones normales, la alfa-sinucleína es una pequeña proteína esencial para el desarrollo del cerebro, ya que tiene una función fundamental en algunos procesos celulares de las neuronas. En condiciones patológicas, la alfa-sinucleína es una de las responsables de la neurodegeneración y la demencia asociada a la enfermedad de Parkinson y las demencias por cuerpos de Lewy. Y todo se debe a la física de la estructura de la proteína y su tendencia a plegarse mal. Voy a visualizarla para enseñartela, con ayuda del software Chimera y los datos del Protein Data Bank.
alfa-sinucleína normal:
Es una proteína sencilla, formada por una cadena en la que se ve, en rojo, una de las formas estructurales fundamentales que forman las proteínas: la alfa-hélice. Dos alfa-hélices se conectan por una cadena simple (un motivo muy básico en las proteíans que se llama hélice-giro-hélice) y queda otra larga cadena simple. En ésta forma, la alfa-sinucleína puede hacer su función fisiológica y es soluble (las alfa-hélices dan lugar a proteínas solubles).
Pero cuando la alfa-sinucleina se pliega mal, algo a lo que esta proteína tiene tendencia debido a su gran simplicidad, tiende a formar otro motivo estructural diferente: la lámina beta. La lámina beta forma estructuras agregadas e insolubles y está favorecida termodinámicamente en estas cadenas cortas de proteína. Las cadenas cortas y simples de proteínas tienden de modo natural a formar fibrillas insolubles. Es decir, el esfuerzo energético para formar la sinucleína funcional de arriba es mucho mayor y, si se la deja a su aire, el fallo en el plegamiento conduce a la:
alfa-sinucleína agregada:
Este es un agregado o fibrilla de alfa-sinucleína. En morado están las láminas beta, que se organizan paralelamente unas con otras, precipitando una masa de proteínas. Forma como «pelusillas» que se van acumulando, agregándose unas a otras, ya que las láminas beta son «pegajosas» y tienden a pegarse unas a otras. La célula marca estas «pelusillas» y las guarda en el agresoma para eliminarlas. Si la célula tiene un defecto para limpiar el agresoma, es cuando se produce la enfermedad.
Cuerpos de Lewy en células de pacientes. B: imagen por microscopía electrónica de un cuerpo de Lewy en el que se observan las masas de «pelusa» de las proteínas agregadas. Mirad la imagen de la estructura anterior e imaginad cómo esas masas se van acumulando hasta formar la «pelusa» de la imagen. La imagen está tomada de éste artículo de Lancet.
¿cómo provocan los cuerpos de Lewy los problemas neurodegenerativos y la demencia?. Pues parece ser que la masa de proteína precipitada interfiere con las funciones celulares, acumula iones de hierro que provocan un fuerte estrés oxidativo que termina matando la célula, dando lugar a la pérdida de las funciones de la zona afectada (motoras, cognitivas) y dando lugar a los síntomas típicos de estas enfermedades.
Al final la demencia por cuerpos de Lewy es una consecuencia natural de la propia Física de las proteínas: la termodinámica nos dice que las cadenas de proteínas, en especial las pequeñas, tienden a formar fibras de láminas beta (como la seda o el nylon). Las células invierten mucha energía y complejos mecanismos para plegar correctamente las proteínas y, cuando el plegamiento falla (por razones termodinámicas o por mutaciones en la cadena que alteran la estructura funcional) hay todo un sistema de control de calidad y eliminación. La enfermedad es el resultado de una cadena de fallos en todo este complejo sistema de construcción, control de calidad y limpieza de las proteínas. Y es el resultado de las propiedades naturales de las cadenas de proteínas, por eso es tan difícil de prevenir y tratar. Resulta extraño pensar que Robin Williams murió por culpa de una pequeña proteína que simplemente se plegó siguiendo su tendencia natural. Si ésto no te hace sentir frágil e indefenso, sometido a las leyes de la Naturaleza, no se que mas podría hacerlo.
Esto nos lleva a otra de las grandes preguntas de la Ciencia: si las cadenas de proteína tienden naturalmente a formar «pelusas» de fibrillas, ¿como se originó, al principio de la evolución, el sistema de estructuras que dió lugar a las proteínas funcionales?