La evolución es un hecho, no un relato. Y las moléculas nos hablan de ello.
escrito por C. Menor-Salvan | 17 diciembre, 2024
C. Menor-Salván 12/2024
Creacionismo y evolucionismo no son creencias o hipótesis alternativas; ni siquiera son conceptos establecidos con las mismas reglas. Son los magisterios no solapados tal como explicaba Stephen Jay-Gould. Así, un supuesto debate o confrontación entre supuestos «científicos» creacionistas y «creyentes en la teoría de la evolución» no tiene sentido.
En primer lugar, ¿de qué hablamos cuando hablamos de «teoría»? La evolución es un fenómeno natural. Por lo tanto, es observable. Sobre un fenómeno natural, los científicos construimos un marco teórico que permite explicar las observaciones y realizar predicciones; éste se actualiza al ir afinando nuestra capacidad para realizar observaciones y experimentos. Pero el hecho ocurre estemos nosotros o no para observarlo y construir hipótesis, modelos o teorías para explicarnoslo.
En todas las teorías hay puntos de especial dificultad. En el marco teórico sobre la evolución tenemos, por ejemplo, la cuestión del origen de la vida, donde observamos que las moléculas de la vida, y sus antecesoras, también siguen reglas de selección, adaptación y supervivencia que aún estamos entendiendo.
No creemos en la evolución; la evolución es un hecho observable
Usando un ejemplo quizá mas fácil de entender: la teoría que explica el funcionamiento de las enzimas también tiene puntos oscuros y aún hay discusión científica en torno a ello. Pero nadie pone en duda que las enzimas existen y su acción es un hecho, sea cual sea la teoría que construyamos para explicarlo. Este marco teórico ha ido cambiando, desde el obsoleto modelo de llave-cerradura hasta modelos como Circe o el de estabilización del estado de transición.
Los científicos no «creemos» en la evolución, del mismo modo que no «creemos» en la enzimas. Son fenómenos reales que tratamos de comprender y explicar. Decir «creo en la evolución» es, simplemente, absurdo.
Como tal hecho observable, las ideas en torno a la evolución no son algo nuevo. Según el historiador romano Diógenes Laercio, Anaxágoras de Clazomene enseñaba que
«los seres vivos se formaron de la humedad, el calor y sustancias terrosas; despues, se propagaron por generación unos de otros».
Esta idea evolutiva rudimentaria, lanzada por Anaximandro de Mileto, el maestro-abuelo de Anaxágoras, hace 2500 años, creaba una disonancia cognitiva con el creacionismo, lo que llevó a San Hipólito de Roma a recogerla en sus «Refutaciones de todas la Herejías«. Gracias a ello, tenemos testimonio de las ideas de la escuela de Anaximandro.
Aún hoy, la evolución es considerada pecaminosa por muchos grupos religiosos. Yo mismo he sido testigo de alguna manifestación en contra de la evolución en EEUU, recibida con bastante humor por parte de los científicos y estudiantes que estábamos en el campus, hay que decir.
Observando la evolución sin salir de tu barrio
La evolución no sólo es observable, sino que la llevamos utilizando siglos en nuestro beneficio. Basta comparar las variedades de lechuga cultivada que podemos encontrar en el supermercado, con su pariente silvestre más cercano y, posiblemente, su ancestro: la amarga e indigesta, aunque comestible lechuga silvestre (Lactuca serriola).
La lechuga silvestre. Antepasado de la lechuga cultivada. Foto: Olga Pokotilo/PlantNet
Todo el proceso de domesticación de plantas y animales llevado a cabo por los humanos durante milenios debería probar, en sí mismo, que la evolución es un hecho. Y no es exclusivo de los humanos. En la evolución de la vida terrestre, unas especies han ejercido presión selectiva sobre otras, condicionando su evolución en una compleja red.
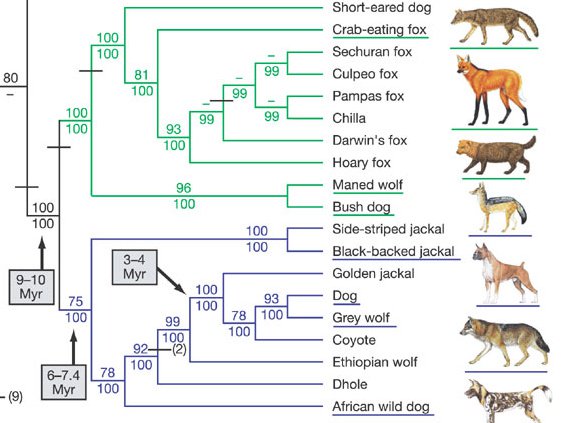
El perro original, que surgió de un ancestro común con el lobo gris actual, se ha diversificado un complejísimo arbol filogenético con todas las razas de perro doméstico. Un ejemplo de evolución divergente impulsada por la presión de selección ejercida por humanos. ¿cual es la frontera entre raza y nueva especie?
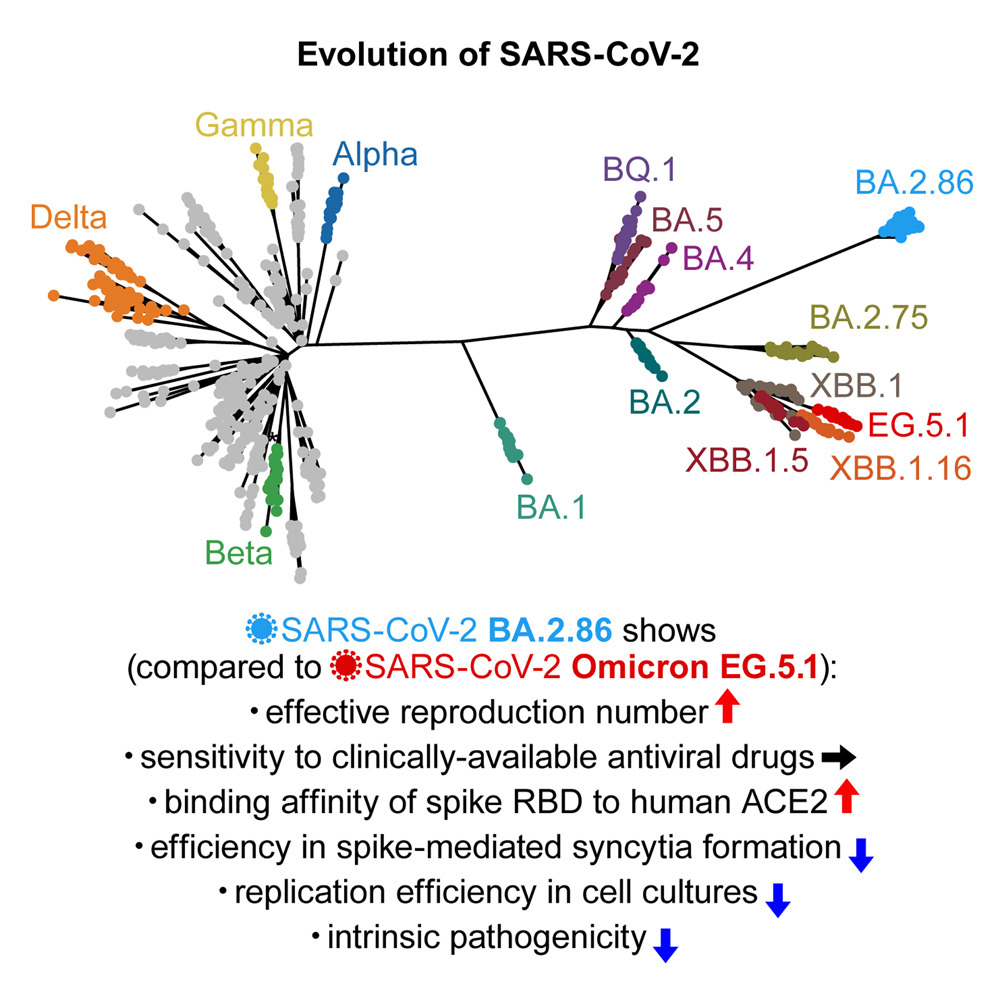
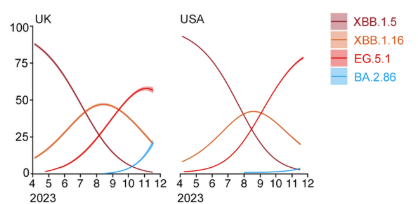
Tenemos un hecho evolutivo aún mas reciente: la pandemia de SARS-CoV-2. Nunca antes se había seguido la evolución de una especie viral tan de cerca y en tiempo real. Desde el inicio de la pandemia, gracias a la velocidad de replicación de los virus, fueron surgiendo variantes, creándose un complejo árbol filogenético. Durante el proceso, muchas variantes se extinguieron y otras han sobrevivido. Actualmente, tanto el virus como la enfermedad COVID son diferentes a los que existían en marzo de 2020. Este proceso de diversificación se basa en pequeños cambios en las estructuras moleculares que ocurren, inevitablemente, durante la replicación. Estos cambios, a veces, son silenciosos, si se mantiene la funcionalidad estructural; pero, a veces, llevan a las variantes a su desaparición, si la funcionalidad de las estructuras disminuye en las condiciones de ese momento, o a su prevalencia, si le aportan una funcionalidad ventajosa en las condiciones del momento. Con el tiempo, puede llegar a surgir una nueva especie.
Arbol filogenético del SARS-CoV-2. Evolución en vivo. Fuente: DOI 10.1016/j.chom.2024.01.001Dinámica de poblaciones de variantes virales. Estas curvas guardan una gran semejanza con las curvas de poblaciones de moléculas en procesos químicos como reacciones metabólicas o evolución química.
Ante todas las evidencias en torno a la evolución de la vida terrestre, la idea del creacionismo y el diseño inteligente se adaptan y, lejos de ser la herejía que añadía San Hipólito, la evolución se vuelve compatible con la idea religiosa de la creación. Así, en el diálogo público, el creacionismo se muestra con frecuencia como un _Deus Ex Machina_ que rellena convenientemente puntos oscuros en nuestra comprensión sobre la vida y el universo, dejando resueltas cuestiones espinosas, como el origen de la vida o del universo. Ello se combina a veces con una visión incorrecta sobre la evolución, cayendo en la trampa teleológica de que conduce hacia la inteligencia como máxima expresión de un proceso perfeccionador, puesto en marcha a partir de unas constantes físicas ajustadas cuidadosamente por un «diseñador» divino con este fin (el principio antrópico fuerte).
Sin embargo, la evolución biológica no es un proceso de mejora dirigido hacia un destino final. Su base es la preservación de estructuras supramoleculares funcionales, que sostienen la continuidad de la vida y que surgieron también mediante un proceso de evolución prebiótica.
La biología molecular es un tratado sobre la evolución
La biología molecular nos ofrece muchos ejemplos para ilustrar la evolución. Dos de los más interesantes son el ribosoma, cuya estructura relata la evolución de la vida desde su origen, y las polimerasas de DNA y RNA, que conectan todos los organismos y cuentan la historia evolutiva de los virus. El problema es que explicar brevemente para el público los complejos detalles moleculares de la evolución no es trivial. Voy a intentarlo, sin embargo, con un ejemplo sencillo (y simplificado), relacionado con un suceso que, desgraciadamente, se repite todos los inviernos: la toxicidad del monóxido de carbono.
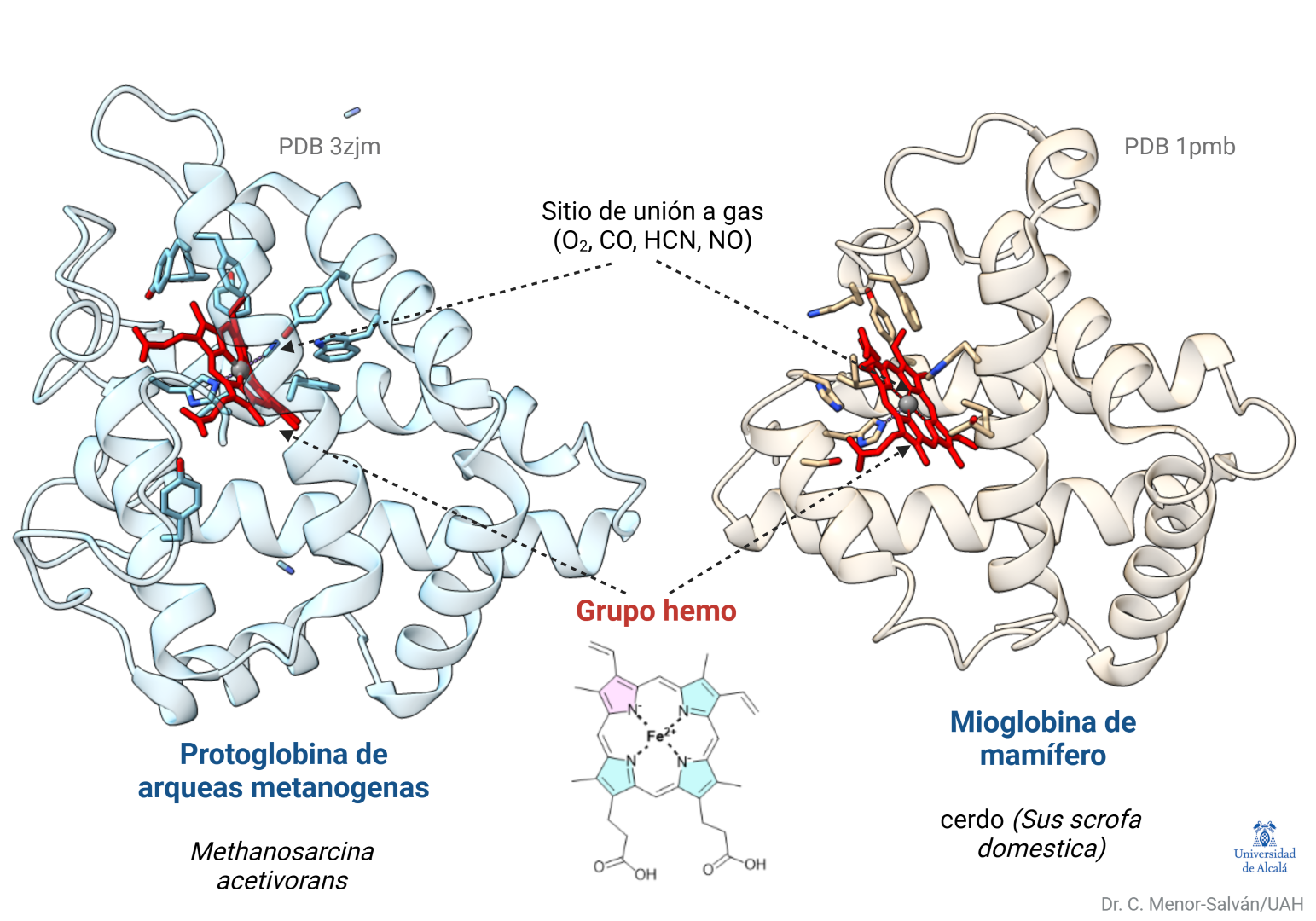
Estructuras de la protoglobina de arqueobacterias y la mioglobina de cerdo, mostrando el grupo hemo en su posición. Ambas proteínas, conectadas evolutivamente, tienen alta similitud estructural y gran afinidad por el monóxido de carbono.
Cuando la vida estaba en sus inicios y la atmósfera no contenía oxígeno, unas arqueobacterias productoras de metano ya estaban dotadas de la proteína protoglobina. Hoy día podemos encontrarlas en lugares tales como sistemas hidrotermales, aguas residuales, algunas minas, donde forman ecosistemas con otros procariotas que usan minerales (sulfuros metálicos) para obtener energía, o en fondos de lagos y mares, donde contribuyen a generar metano atmosférico.
Estas arqueas usan un sistema ancestral para metabolizar monóxido de carbono y usarlo como fuente de energía y carbono. En los inicios de la vida, posiblemente la protoglobina era un sensor de monóxido de carbono y, tal vez, de cianuro, otro componente quizá presente en aquel ambiente. Ambos gases se unen fuertemente al grupo hemo de la protoglobina.
Nosotros tenemos mioglobina y hemoglobina, que almacenan y transportan oxígeno. Estas proteínas evolucionaron a partir de aquellas globinas ancestrales de las arqueas, de quienes heredamos muchas estructuras moleculares, pues somos el resultado de una unión entre arqueas y bacterias que ocurrió hace unos 2000 millones de años.
Hace unos 600 millones de años, comenzó la evolución de la hemoglobina, que posibilitó el transporte de oxígeno desde el ambiente a los órganos, favoreciendo la evolución de los animales. Una consecuencia de este proceso de evolución es que el monóxido de carbono es muy tóxico para nosotros.
Esta toxicidad se debe a que nuestras mioglobina y hemoglobina tienen mucha afinidad por el monóxido de carbono, como la protoglobina arqueal; quizá, es un recuerdo de aquel lejano ancestro, que vivía en un ambiente sin oxígeno, alimentándose del monóxido de carbono. La mioglobina y hemoglobina no son el resultado de un «diseño inteligente», sino que resultan de la evolución de algo que surgió en un ambiente sin oxígeno y cuya función original se convirtió en un problema millones de años después.
Y no es el único problema. La vida nació en un ambiente sin oxígeno; cuando este empezó a aumentar en la atmósfera, tuvo lugar una de las primeras grandes extinciones. Los supervivientes desarrollaron adaptaciones moleculares que «parcheaban» los problemas que surgieron. En un ambiente sin oxígeno, el hierro reducido de la protoglobina era estable; en el ambiente con oxígeno, el hierro de la mioglobina y hemoglobina se oxida, por lo que tuvo que evolucionar un sistema antioxidante.
Otro de estos parches es una modificación en nuestras globinas que reduce ligeramente su afinidad por el monóxido de carbono. Ello reduce lo suficiente la toxicidad del gas como para que los fumadores puedan dar las gracias por ello. Sin esa modificación, los animales no soportaríamos un poco de humo.
La histidina 64, presente en la hemoglobina pero no en la protoglobina, estabiliza la unión del oxígeno al grupo hemo de la hemoglobina, que se produce formando un ángulo, mientras que obstaculiza ligeramente la unión del monóxido de carbono. Aunque el CO sigue teniendo mucha más afinidad por la hemoglobina que el O2, este pequeño cambio aportó una ligera ventaja a los animales.
Este ejemplo ilustra una característica básica de la evolución: el mantenimiento y adaptación de estructuras funcionales, no la mejora hacia un diseño óptimo. La mioglobina de mamífero y la protoglobina de arqueas sólo tienen en común un 13% de la secuencia de sus genes. Ese pequeño porcentaje permite que sus estructuras y función básica (la unión de estos gases) se hayan preservado y guardan una gran similitud estructural.
Las moléculas biológicas nos conectan a todos los organismos en un gran árbol cuya raíz está en el origen de la vida, hace más de 4000 millones de años. Y esto es un hecho, no un relato.
¿Estamos solos en la galaxia?
escrito por C. Menor-Salvan | 17 diciembre, 2024
C. Menor-Salván. Agosto 2023
*ver The Conversation para una versión divulgativa corta más actual.
«»A veces creo que hay vida en otros planetas, y a veces creo que no. En cualquiera de los dos casos la conclusión es asombrosa.»»
— Carl Sagan
Esta cuestión lleva resonando en nuestra mente colectiva desde que los humanos comenzaron a observar el Cosmos desde un punto de vista científico, sin invocar dioses ni mitos. Descubrir que no estamos solos en nuestra galaxia o en el Universo sería el mayor y más relevante hallazgo de la Historia humana. Pero, por el momento, solo podemos especular, con más o menos fundamento, acerca de ello.
La pregunta ha vuelto a ponerse de moda a raíz de la reciente declaración de unos ex-militares estadounidenses encabezados por el oficial de la USAF David Grusch, que aseguran que la fuerza aérea de los EEUU oculta en secreto naves extraterrestres y ‘restos biológicos no humanos’ (¿plantas? ¿animales? ¿alienígenas?). Para contextualizar, aquí me entrevistaron sobre ello.
No vamos a hacer una discusión académica sobre la posible vida extraterrestre y la paradoja de Fermi, sino tan sólo delimitar la cuestión (u organizar nuestra ignorancia), hacer algunas reflexiones sencillas al alcance de cualquier lector y dar mi opinión como astrobiólogo de estas ‘nuevas’ (veremos que de nuevas nada) declaraciones. En las referencias se encuentra la bibliografía que he usado y que permitirá al lector interesado profundizar en el tema.
En este artículo, vamos a seguir esta línea de razonamiento:
Acotando la terminología: ‘vida’ vs ‘vida inteligente’
La vida terrestre como modelo, el único que tenemos: cuándo surge la vida y cuándo surge la ‘vida inteligente’.
La vida puede ser común, pero la inteligencia no tiene por qué surgir siempre.
¿cómo buscamos vida extraterrestre?: Sistema Solar y exoplanetas.
¿cómo buscamos vida inteligente? escuchando señales. Una estrategia condenada al fracaso: la galaxia (no ya el universo) es muy vasta. Si están puede que no los escuchemos nunca, ni ellos a nosotros.
La tecnología, ¿podrá resolverlo en el futuro o nos acercamos al final del desarrollo tecnológico?
¿nos han visitado alienígenas?. ¿hay evidencias? ¿es posible una galaxia tipo ‘Star Trek’?
Conclusiones
A qué nos referimos cuando decimos ‘solos en el universo’
Cuando los científicos hablamos de vida extraterrestre no nos estamos refiriendo a ‘vida inteligente’ sino a cualquier forma de vida. Nos interesa descubrir cómo se origina la vida, cómo evoluciona en un planeta, si la vida se originó en otro lugar además de en la Tierra, o si hay lugares que reúnen las condiciones para que surja vida o que se encuentran en la fase de evolución química (lo llamamos ‘habitabilidad’). Todos ellos son parte de los objetivos de la ciencia de la Astrobiología.
Esquema general de la evolución de la vida terrestre y de las aproximaciones para entender su origen.
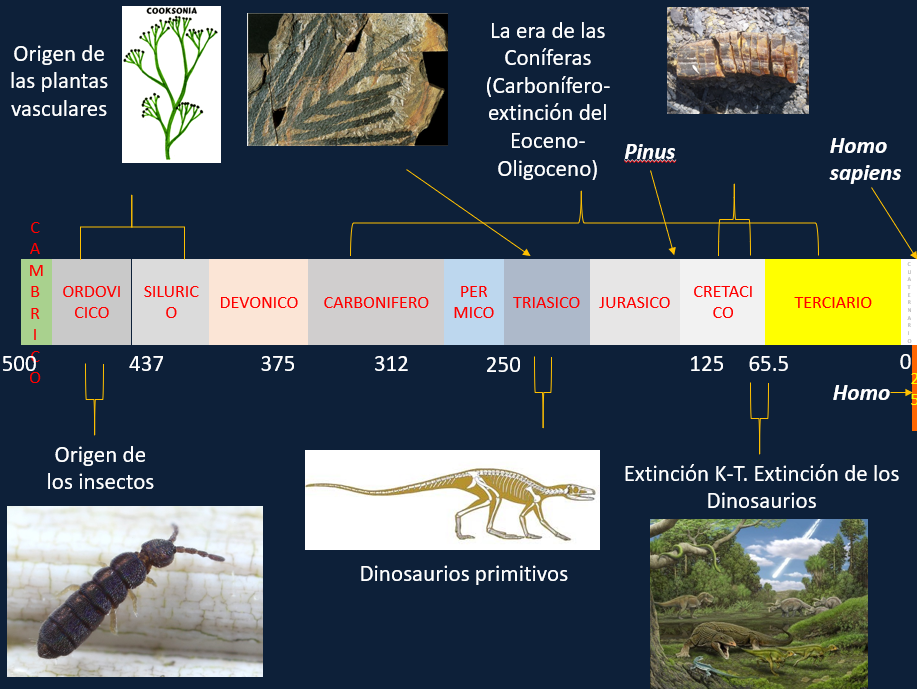
Usando nuestro planeta como referencia, hay que tener en cuenta que calculamos que la vida se originó aquí hace unos 4200 millones de años. Se calcula que la vida pudo surgir en unos 40 a 120 millones de años desde que comenzó a ser habitable, lo cual es bastante rápido en términos geológicos. Probablemente, una vez dados los primeros pasos, el proceso debió ser muy rápido.
Durante siguientes 2500-3000 millones de años, la vida terrestre estuvo dominada por organismos unicelulares o pluricelulares muy simples (antecesores de las algas actuales). Los animales habitan la Tierra desde hace unos 500 a 600 millones de años. Es decir, los animales han ocupado el planeta durante el último 14% del tiempo desde que existe la vida en éste. Prácticamente somos unos recién llegados.
Durante los primeros 2000 millones de años de vida en la Tierra la situación era mas o menos así: cianobacterias. Si encontrásemos vida extraterrestre, lo más probable es que se parezca más a esto que a ET o Predator.
Dado que el concepto de inteligencia, como el de vida, es algo difuso, definiremos como ‘vida inteligente’ aquella con la capacidad de observar el cosmos.
¿cuándo surgió en la Tierra una forma de vida que comenzó a poner sus ojos en el firmamento y a andar el camino hacia su exploración?.
Si ponemos la Edad de Bronce como el inicio de las primeras observaciones astronómicas sistemáticas, digamos que tenemos vida inteligente en la Tierra capaz de observar el Cosmosdurante el último 0.00000012% del tiempo de existencia de vida. Los científicos también hablan de vida inteligente con tecnología de comunicación capaz de mandar señales fuera del planeta. Los humanos somos capaces de ello desde la invención de la radio, es decir, desde hace poco más de un siglo.
Cálculos recientes, basados en el análisis bayesiano de los datos de la vida que conocemos, sugieren que hay aproximadamente un 60% de probabilidades de que nunca surja vida inteligente en un planeta con vida. Es decir, la evolución de la vida no necesariamente va a conducir a que emerja vida inteligente. Si la vida es una consecuencia de la evolución del cosmos, no tenemos razones para pensar que la inteligencia también lo sea.
Con estos datos, es fácil asumir que, si encontramos evidencias de vida extraterrestre, seguramente serán de vida simple (unicelular o pluricelular) o, en general, vida no inteligente. Como es lógico, no necesariamente tiene que ser así, pues no conocemos cuánto tiempo puede tardar en emerger vida inteligente en un planeta; en la Tierra puede que haya tardado mucho, dada la cantidad de eventos de extinción que han ocurrido.
O bien, al contrario, puede que el impulso evolutivo que dan las crisis biológicas haya hecho que surja más pronto. No hay forma de saberlo aún. Pero, con lo que conocemos, no es descabellado asumir que, probablemente, la vida sea frecuente en la galaxia, pero que la vida inteligente sea bastante poco común o incluso única. De hecho, otros cálculos recientes sugieren que, en nuestra galaxia, podría haber entre 1 y 10 planetas con vida inteligente capaz de explorar el cosmos, incluyendo el nuestro.
Los últimos 500 millones de años de vida en la Tierra son los más animados desde el punto de vista biológico. La barra está a escala; obsérvese cuando surgen los humanos.
En este sentido, hay que aclarar que la ecuación de Drake (formulada a partir de una famosa pregunta de Fermi a sus estudiantes: ¿cuantos pianos hay en Chicago?) no pretende ser una estimación precisa, sino una aproximación mental y una delimitación de nuestra ignorancia. El propio Drake solía responder que el número de civilizaciones está «entre una y mil millones».
Entonces, ¿no tenemos evidencias de vida extraterrestre?
No, no tenemos ninguna evidencia. Buscamos evidencias de vida, es decir, biofirmas, de varias formas:
Huellas moleculares, geoquímicas o físicas. Estas pueden ser biomarcadores orgánicos o inorgánicos, es decir, compuestos químicos que estén ligados a la vida. Pueden ser huellas geoquímicas o geológicas, tales como determinados minerales o huellas isotópicas.
También pueden ser señales espectroscópicas o biofirmas basadas en fenómenos químico-físicos, tales como desequilibrios químicos, estructuras termodinámicas lejos del equilibrio.
O, simplemente, pueden ser indicios de habitabilidad, tales como presencia de agua líquida o temperaturas en el rango de estabilidad de moléculas orgánicas
Buscamos todas estas biofirmas, bien de vida o bien de evolución química, e indicios de habitabilidad en otros lugares de nuestro Sistema Solar que pudieron ser habitables (Marte) y en los exoplanetas, planetas descubiertos en otros sistemas planetarios.
La búsqueda de biofirmas en exoplanetas aún es un campo en el que hay mucho que desarrollar y se basará fundamentalmente en la búsqueda de señales espectroscópicas que indiquen habitabilidad o vida y que nos permitan conocer la composición atmosférica en cierto detalle.
Los astrónomos están investigando exhaustivamente las estrellas que nos rodean, nuestro vecindario cósmico, buscando nuevos planetas. El catálogo de exoplanetas de la ESA y el catálogo de la NASA indican que, mientras escribo estas líneas, se han identificado nada menos que 5483 planetas en 4220 sistemas planetarios, todos en nuestra vecindad galáctica. Esta lista incluye planetas desde tamaños mucho mayores que Júpiter a tamaños similares e incluso menores a la Tierra. Algunos eran prometedores candidatos a ser planeta habitable.
Sin embargo, al profundizar en su observación, todavía no hemos encontrado ningún planeta que sea un serio candidato a reunir condiciones de habitabilidad o vida. En los planetas cuya temperatura han podido estimar los científicos, todos salvo uno son demasiado calientes para sostener vida orgánica. Y el único con temperatura moderada, es un gigante gaseoso mayor que Júpiter.
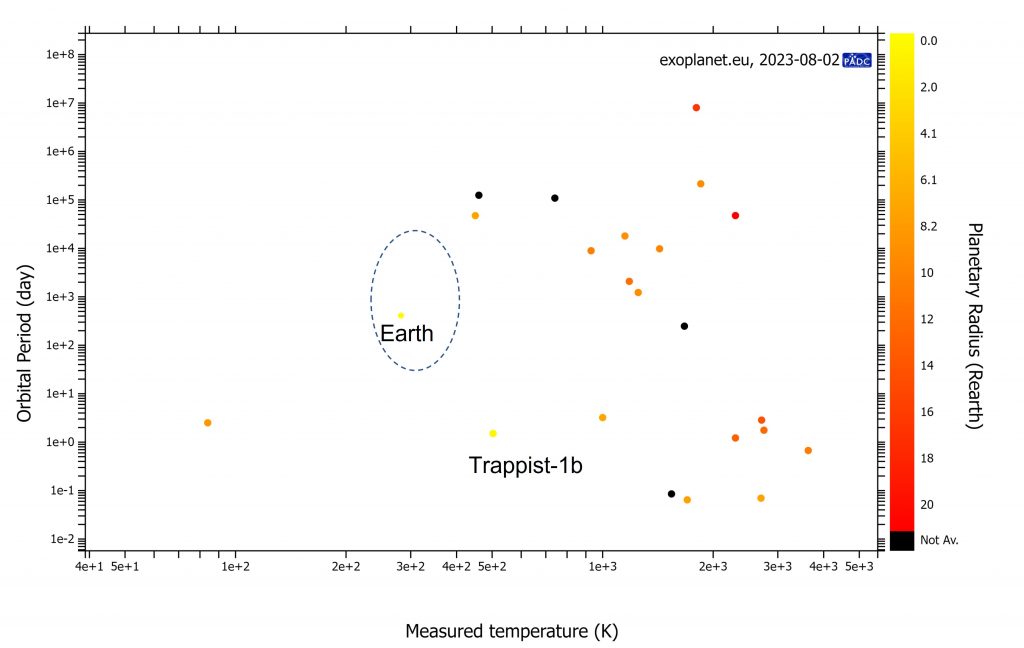
Scatter plot de los exoplanetas cuya temperatura ha podido determinarse hasta la fecha de hoy. La temperatura mas baja, 84K, se corresponde al planeta Qatar-3b, situado en el sistema Qatar-3, una estrella similar a nuestro Sol situada a más de 2300 años luz. El planeta es un enorme ‘Júpiter caliente’ y la estrella es demasiado joven (0.3 Ga), así que la vida está prácticamente descartada en ese sistema. Hemos ubicado el lugar donde estaría la Tierra en este gráfico y la región donde esperamos planetas con posible vida (circulo). Trappist-1b y 1c, a 40 años luz de la Tierra, fueron muy prometedores cuando se descubrieron; la medida de su temperatura da un valor medio de 503K, demasiado cálido para sostener vida orgánica, además de estar en bloqueo de marea y la proximidad a su estrella, lo que sugiere alta irradiación. Trappist-1b tiene una atmósfera densa rica en agua, lo que sugiere que en el pasado pudo tener océanos y ahora es demasiado caliente (efecto invernadero desbocado).
La mayor parte de los exoplanetas identificados son demasiado calientes, sometidos a intensa radiación, están en acoplamiento de marea o, en general, tienen características que descartan su habitabilidad; pero, la mayoría de los sistemas planetarios tienen edades comparables al nuestro y muchos tienen múltiples planetas. La presencia de planetas habitables (y no digamos habitados) no parece demasiado común, al menos de momento y en nuestro vecindario cósmico, pero no perdemos la esperanza y, posiblemente, con la mejora de la tecnología de observación tengamos sorpresas pronto.
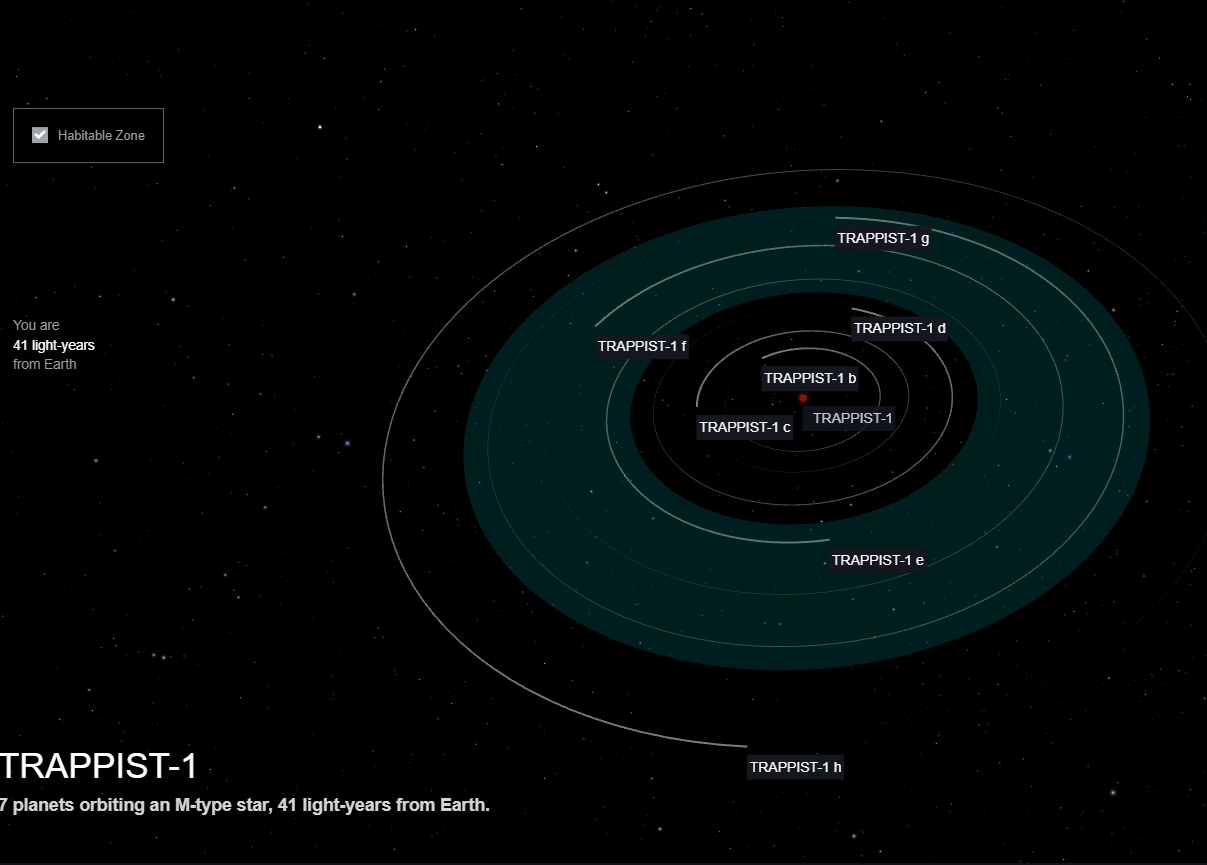
El sistema Trappist-1, a tan sólo 41 años luz de nosotros, contiene 7 planetas, varios de ellos rocosos y de tamaños comparables a la Tierra. Su descubrimiento causó gran agitación: ¿se encontró un sistema planetario con posible vida?. Varios de sus planetas están en la región de habitabilidad. Sin embargo, su estudio detallado sugiere que ninguno de los planetas es habitable, aunque todavía no hay nada concluyente. Se ha sugerido que la estrella tiene una edad avanzada y que varios de sus planetas, como el 1b y 1c, pudieron ser habitables en el pasado. Actualmente, 1f podría ser un mundo helado similar a Europa o Encelado. No obstante, no hay nada cerrado y futuras observaciones pueden darnos sorpresas.
Pero, por el momento, no hay evidencias de vida extraterrestre y ni siquiera podemos confirmar que ninguno de los planetas observados sea habitable en el sentido astrobiológico. Ya podemos ir haciendo números con lo que sabemos: se han observado unos 1800 planetas rocosos (tipo terrestre o ‘supertierras); aunque hayan surgido sospechosos, no podemos confirmar que haya vida o condiciones de habitabilidad en ninguno de ellos.
Ello implica que, de momento, hay una probabilidad de un 99.94% de que un planeta rocoso tipo terrestre recién encontrado no tenga vida. Encontrar un planeta habitable es algo parecido a que te toque la lotería. La diferencia es que nuestra galaxia juega muchos números.
Pero, no sólo debemos tener en cuenta los planetas. Se han observado 849 planetas con masa mayor que la de júpiter. Estos planetas pueden tener lunas con posibilidades de habitabilidad, como, en nuestro sistema solar, es el caso de Europa o Encelado. Aún no hay capacidad tecnológica para observar las exolunas, aunque ya hay algún candidato a exoplaneta con sistema de lunas. Ello multiplica las posibilidades de que exista vida en nuestro entorno planetario.
Impresión de cómo podría ser la exoluna Kepler-1625b-i, que orbita el planeta Kepler-1625b, un ‘superjúpiter’ de aproximadamente el tamaño de nuestro júpiter pero mucho más denso. Confirmar la existencia de exolunas es muy difícil por el momento.
Por ello, debemos seguir investigando, pues queda mucho espacio por observar y aún tienen que mejorar mucho las observaciones. Puede que en un futuro próximo alguno de los observados, o uno nuevo, pueda confirmarse adecuado para el origen o presencia de la vida.
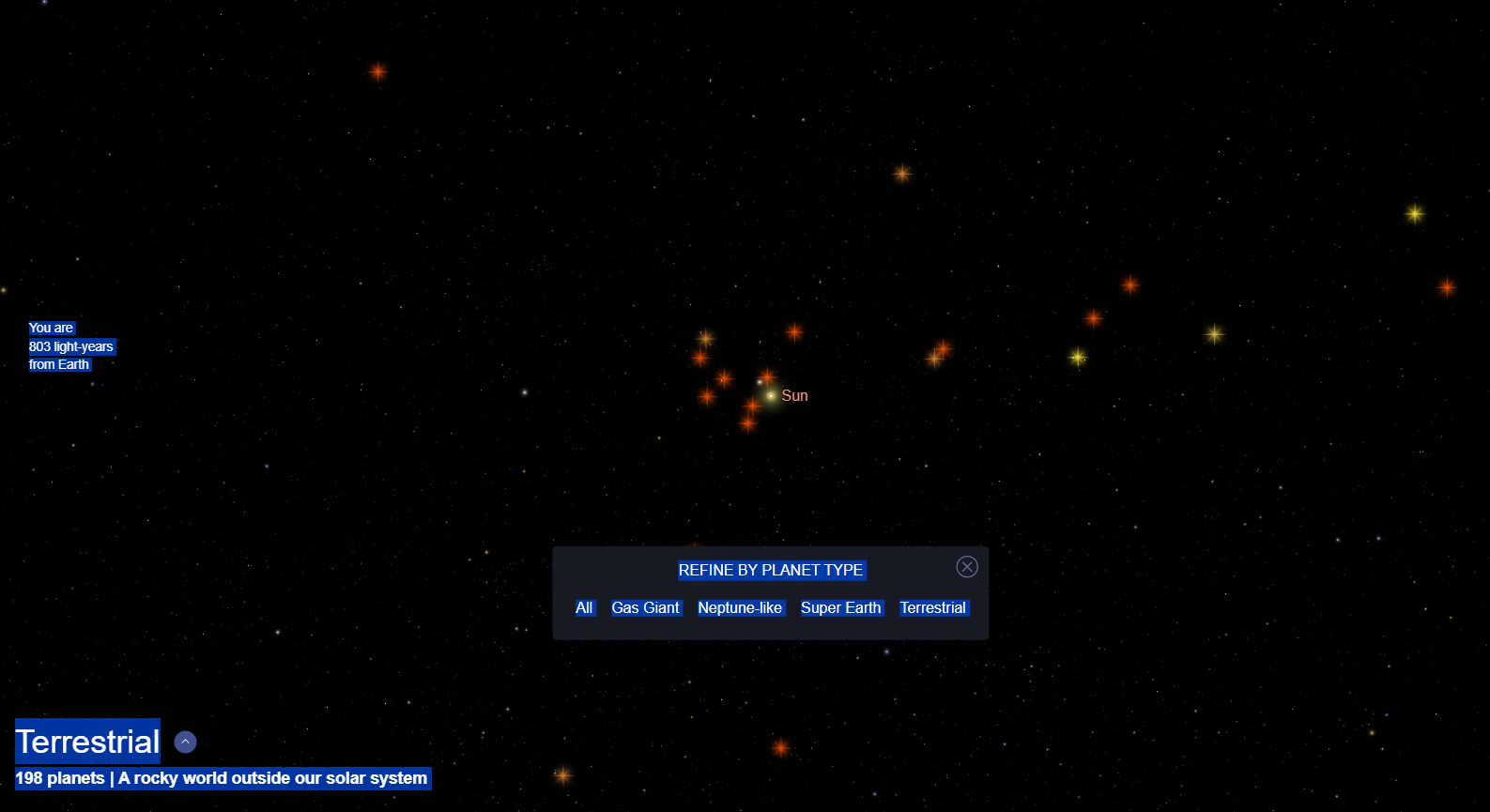
Mapa en 3D de los sistemas con planetas de tipo terrestre (rocosos y tamaños similares a la Tierra) descubiertos por ahora en nuestro vecindario galáctico, en el precioso catálogo de exoplanetas de la NASA. De todos los planetas identificados, 198 son de tipo terrestre pero ninguno de ellos parece ser habitable. De momento no hay vulcanos ni klingons ahí fuera.
¿la vida podría ser frecuente en la galaxia o no?
Pensamos que la formación de vida puede ser relativamente común por varias razones: la universalidad de la química prebiótica, que da lugar a la formación de un set similar de compuestos orgánicos en muchas condiciones y que hemos observado en asteroides, por ejemplo. Es decir, la formación de los precursores orgánicos simples de la vida es un proceso que se integra dentro de la geoquímica de modo normal y, seguramente, es muy corriente en la galaxia.
Por tanto, el primer paso hacia la vida orgánica debe ser común. Tal vez el siguiente paso, el proceso de evolución química, sea menos común. Pero no tenemos motivos para pensar que sea una rareza, ya que es un proceso que se integra en la geoquímica planetaria.
Por otro lado, la evolución de la vida desde un punto de vista termodinámico parece una consecuencia de la propia evolución de los sistemas planetarios. La vida es un eficiente sistema de generación y disipación de entropía. La entropía del universo aumenta, la energía de las estrellas se disipa y, en el proceso, surgen estructuras que podrían maximizar la producción de entropía. La vida surgiría, entonces, como una estructura dinámica en el conjunto de fluctuaciones generadas por los procesos de disipación de energía y aumento de entropía del universo, es decir, una consecuencia del segundo principio de la termodinámica.
Pero, si la evolución y vida parece un imperativo de la propia termodinámica del universo, entonces, ¿por qué todavía no hemos visto ningún planeta habitable o con pistas de vida?.
Por varias razones: aunque la vida sea probable, hemos observado sólo una muestra mínima de planetas. Se calcula que solo en nuestra galaxia hay unos 100.000 millones de planetas. En nuestro vecindario, hasta 50 años luz de distancia, hay unos 1500 planetas; se han observado aproximadamente el 10% de ellos. Es muy probable que la vida sea frecuente en números absolutos, aunque la probabilidad de encontrar un planeta con vida o habitable sea baja; tan sólo hemos observado el 0.000005% de los planetas de nuestra galaxia.
No puede decirse ni que tengamos mala suerte buscando, pues ni siquiera conocemos al 90% de los vecinos de nuestra propia manzana.
Si nosotros no los vemos, ¿pueden vernos ellos?
Si hay un planeta con vida inteligente en un radio de unos 3000 años luz, con una tecnología de observación similar o superior a la nuestra, cabría la posibilidad de que hubieran encontrado nuestro Sistema Solar y catalogado nuestro planeta como habitable o con vida, en base a las huellas espectroscópicas, distancia del Sol, masa, movimiento y otras características.
Esto podría ocurrir ahora (lo cual es extremadamente improbable), o haber ocurrido en algún momento durante los últimos cientos o miles de millones de años. O podría ocurrir en un futuro. No hay nada que descarte que nuestro planeta ha sido ya observado.
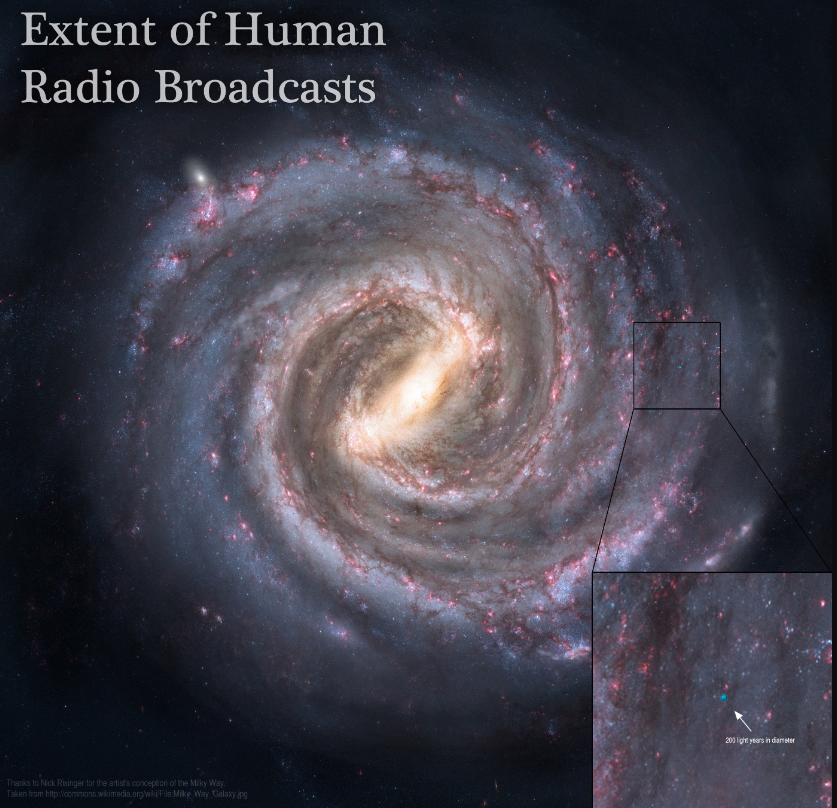
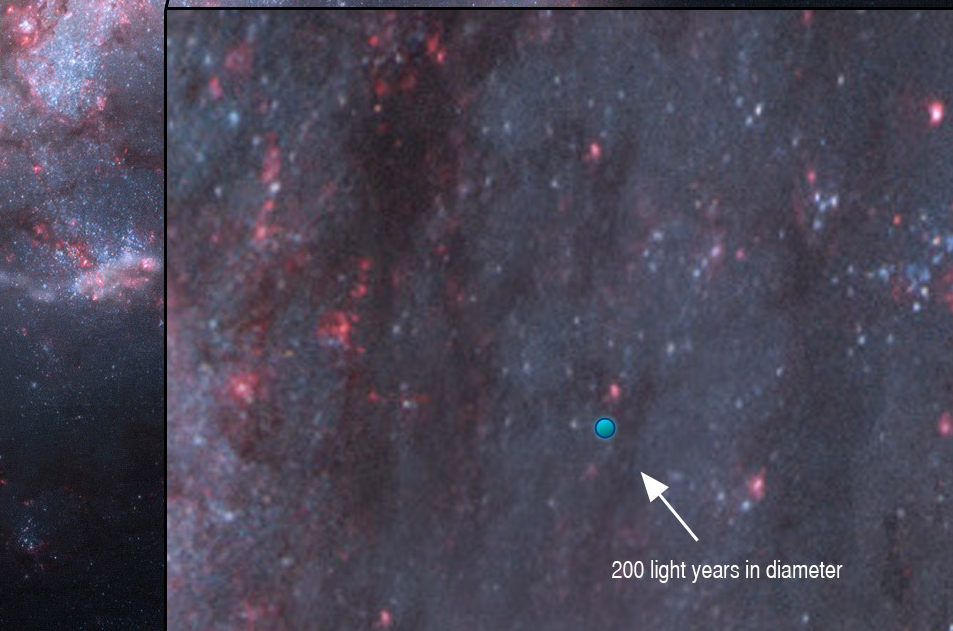
En cuanto a detectar nuestras emisiones de radio y determinar que provienen de seres inteligentes, es mucho menos probable. Teniendo en cuenta que llevamos emitiendo señales al espacio unos pocos años, estas serían detectables en una pequeña burbuja de unos 200 años luz de diámetro. A través del proyecto SETI, los humanos llevan varios años buscando débiles señales que sugieran vida inteligente. Nunca se recibió ninguna. Una vez hubo un susto, la famosa señal wow, pero no hubo suerte.
No obstante, llevamos poco tiempo escuchando. Sería como encender la radio durante un segundo, justo cuando no se oye a nadie hablando, y pensar que no hay nadie ahí. O, como nos ocurre a nosotros, la extensión de la burbuja de emisiones de una civilización extraterrestre quizá quede demasiado lejos. Es como tratar de comunicarnos con alguien usando un walkie talkie o una pequeña emisora de poco alcance. Si no hay nadie con un receptor, nadie va a oir nuestra emisión ni responder. Pero ello no implica que no haya nadie en los alrededores.
Extensión de nuestras emisiones de señales: el pequeño punto azul, insignificante en la galaxia. Imagen de Adam Grossman y Nick Risinger, que puede verse en alta resolución, aqui en The Planetary Society.Detalle de la burbuja azul de nuestras emisiones de radio. Detectar emisiones de seres extraterrestres implicaría que su burbuja nos ha alcanzado. Esto es muy poco probable, aunque hayan estado emitiendo.
Además hay que tener en cuenta que las señales electromagnéticas (radio, luz,…) disminuyen su intensidad de modo proporcional a 1/d2, con lo que necesitan instrumentación sensible. La probabilidad de que haya una civilización o inteligencia extraterrestre buscando señales extraterrestres (para ellos) con instrumentación sensible, en un área conteniendo unos 6000 planetas, es ínfima, máxime cuando los cálculos más optimistas sugieren que, en este momento, la galaxia puede contener unos 10 planetas con vida inteligente capaz de explorar el cosmos.
Así que, quizá estén ahí fuera y, ni nos oigan, ni les oigamos jamás. O quizá surja en el futuro una civilización que detecte una débil señal procedente de nuestro planeta, donde una civilización desapareció miles de años antes.
Quizá sea una cuestión de limitación tecnológica: el gran filtro
Aquí la imaginación es el límite: podemos imaginar que una inteligencia extraterrestre ha encontrado formas de escuchar señales lejanas o viajar a otros puntos de la galaxia. Pero lo que podamos imaginar no nos sirve si no lo podemos sustanciar en una aproximación científica.
Podemos afirmar que quién sabe qué tecnologías desarrollaremos. Este pensamiento es muy común: «hace 100 años apenas éramos capaces de volar, y ahora salimos al espacio. ¡Quién sabe qué haremos en 100 años más!»
Esta idea presupone una extrapolación lineal en el desarrollo de la tecnología. La mente humana tiende a pensar de modo lineal: si en 100 años hemos conseguido tal cosa, en otros 100 extrapolamos esto linealmente y llegamos al mundo de Star Trek.
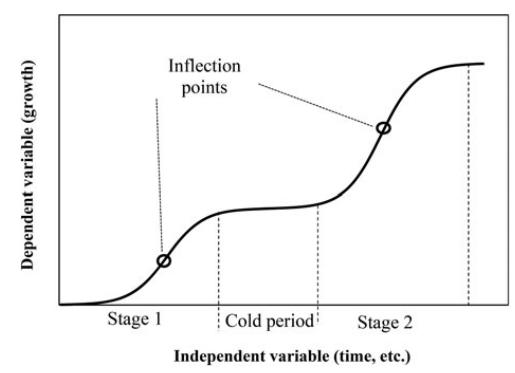
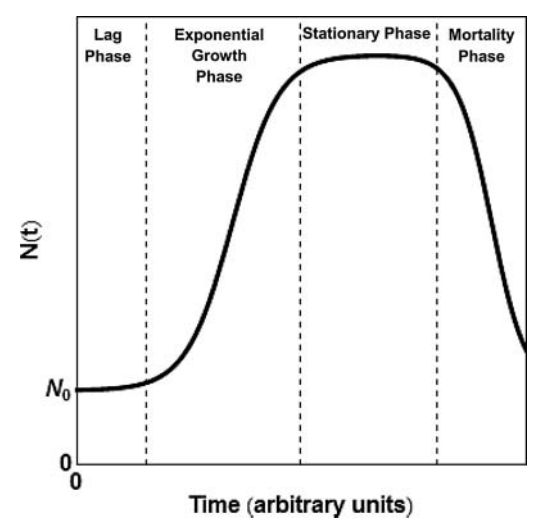
El problema es que la Naturaleza no suele seguir funciones lineales. El desarrollo tecnológico humano, como tantos procesos de desarrollo o crecimiento, sigue una función sigmoidal. Voy a usar como analogía el crecimiento de la colza:
El conocimiento y la tecnología siguen un patrón similar. Tras una fase inicial lenta, se llega a una fase de aceleración exponencial, seguida de una fase de meseta, en la que el rápido incremento en el conocimiento y la tecnología se frena. Aquí pueden ocurrir dos cosas: que termine declinando la civilización y se pierda el conocimiento y la tecnología, o que haya un ‘salto evolutivo’, una nueva tecnología o cambios de paradigmas que permitan un nuevo periodo de crecimiento. ¿Dónde estamos nosotros ahora?. ¿llegando a un punto de inflexión? ¿entrando en un periodo frío? No se si alguien lo habrá estudiado, pero no he encontrado referencias. Así que, teniendo en cuenta las funciones sigmoidales, dado que hemos tenido un rápido crecimiento tecnológico y en conocimientos, en algún momento se llegará a una meseta, si no estamos llegando ya.
Teniendo en cuenta que todos los fenómenos de crecimiento y poblacionales siguen una función sigmoidal (ya sea el crecimiento de una colonia de bacterias, de un bosque, de la capacidad de computación, de una población, de una pandemia, de la economía….), la idea del ‘gran filtro de Hanson‘ es consistente con un modelo sigmoidal simple: podemos sugerir que toda civilización inteligente llega, tras un periodo de rápido crecimiento tecnológico, a una meseta, seguida de decaimiento y, eventualmente, desaparición de esa civilización. Dado que es una ley natural de los procesos de desarrollo poblacionales, con seguridad una civilización extraterrestre basada en poblaciones de individuos va a seguir ecuaciones sigmoidales.
Todo crecimiento en un sistema cerrado (ya sean bacterias en un cultivo o una civilización) siguen una curva sigmoidal que llega a una meseta y decaimiento (extinción). Este modelo es consistente con la idea del ‘gran filtro’
La idea del gran filtro implicaría que, en un planeta, como sistema semiabierto (como es por ejemplo un cultivo bacteriano), toda civilización llegaría a la fase de extinción antes de que tengamos la posibilidad de encontrarnos. La única forma de evitarla (como en un cultivo bacteriano) es contactar con el exterior y ‘pasar’ parte de la población a un nuevo sistema (pasando bacterias a un nuevo cultivo). Entonces, si no existe la posibilidad física de sortear la distancia o de encontrar ninguna otra de esas civilizaciones o inteligencias potenciales, no nos queda otra que extinguirnos en silencio. Un mundo como el de Star Trek resolvería el problema del gran filtro; implicaría que hay un salto tecnológico, de conocimiento o evolutivo que no podemos imaginar y que nos llevaría a un nuevo periodo de crecimiento exponencial, hasta el siguiente punto de inflexión.
En Star Trek: First Contact, el hallazgo del imaginario ‘motor de curvatura’ y la llegada de los vulcanos salva a los humanos de caer en el gran filtro.
Entonces, ¿nos ha visitado una inteligencia o inteligencias extraterrestres? ¿qué hay de los recientes testimonios sobre alienígenas en el Congreso de los EEUU?
Las noticias y testimonios sobre el fenómeno OVNI y visitas extraterrestres han existido desde el siglo XIX. Periódicamente, el mito de los misteriosos hangares y centros de investigación estadounidenses con naves extraterrestres y restos de alienígenas se reaviva. Surgen oscuros testimonios, vagas explicaciones, imágenes borrosas que sirven, sobre todo, para hacer películas de ciencia ficción sobre el tema.
Pero, no hay ni una sola prueba.
Los científicos no trabajamos con anécdotas ni testimonios. Trabajamos con evidencias. Usamos la ‘Navaja de Hitchens’:
una afirmación sin evidencias se puede descartar sin evidencias»
Los científicos (los periodistas, en teoría, también. Christopher Hitchens era periodista) simplemente descartamos los testimonios que no se sustentan en pruebas, datos y evidencias. Y en afirmaciones como esta se requieren datos sólidos. Como dijo Carl Sagan, afirmaciones extraordinarias requieren pruebas extraordinarias.
David Grusch, uno de los declarantes en el Congreso sobre los UAP y la supuesta tenencia de tecnología y restos biológicos extraterrestres. Una declaración ridícula, hueca y, desde el punto de vista científico, totalmente descartable. Personalmente, me pareció todo una broma.
Estas recientes declaraciones en el congreso de EEUU han sido un compendio de los clásicos testimonios sobre fenómenos OVNI que hemos visto en otras ocasiones: nada concreto, ninguna prueba, evasivas e incluso afirmaciones, digamos, interesantes, como que el declarante sugiere que los extraterrestres estaban muy interesados en la tecnología nuclear. ¿una inteligencia con tecnología tan avanzada como para cruzar cientos o miles de años luz, interesada en una tecnología seguramente obsoleta? ¿tal vez quiere decir que la energía nuclear es muy importante y en su planeta se quedaron sin uranio?.
El testimonio era incluso ridículo cuando le preguntan al declarante si había visto personalmente esos restos de naves alienígenas y restos biológicos ‘no humanos’ y dice que no personalmente, que se lo han contado. No sólo declaran sin aportar pruebas, sino en base a rumores.
El ‘black mailbox’, en Nevada. En la ruta de los entusiastas del ‘fenómeno OVNI’, próxima al área 51. Aquí esperan ver o recibir alguna comunicación de los extraterrestres.
Estas declaraciones ni siquiera pueden considerarse ciencia ficción, sino meras elucubraciones y rumores. Además, estas cosas no tienen en cuenta algo elemental sobre la naturaleza humana: no sabemos guardar secretos. ¿un centro de investigación con naves extraterrestres capturadas, lo que sería el hallazgo más importante de la Historia de la Humanidad, por el que habrían pasado miles de trabajadores, científicos y militares, oculto y secreto desde los años 1930?.
Este nuevo episodio de declaraciones sobre supuestas tecnologías extraterrestres me recuerda al caso de Bob Lazar. Este curiosísimo personaje afirmó haber trabajado en el ‘área 51’ y en un lugar secreto llamado S-4, donde se realiza ingeniería reversa de naves extraterrestres, y realizó declaraciones muy similares a las que se han escuchado estos días en el Congreso de los EEUU. Afirmó que unos seres extraterrestres provenientes de Zeta-Reticuli, un sistema estelar situado a unos 39 años luz, utilizaban en sus naves el ‘elemento 115’, de propiedades muy inusuales. Bob Lazar llegó a ser muy popular en el colectivo de los conspiranoicos y fans del fenómeno OVNI. Sin embargo, el descubrimiento de que zeta-reticuli carece de planetas en su sistema y la síntesis del elemento 115 en 2003 demostraron que las afirmaciones de Bob Lazar no eran más que o delirios, o bien un plan para ganar notoriedad y hacer su negocio en torno al mundo OVNI.
Bob Lazar con uno de sus esquemas de naves extraterrestres. ¿embaucador, perturbado o visionario?
Bob Lazar es un personaje curioso no obstante. Siempre me preguntaré de dónde obtuvo muestras de plutonio (cantidad ínfima, obviamente), de las cuales estuve a punto de adquirir una para mi colección cuando fue detenido por el FBI.
Los científicos debemos mantener la mente abierta. Debemos investigar. Pero investigamos siguiendo una serie de metodologías que evitan que caigamos en las trampas cognitivas comunes o terminemos cayendo en las redes de embaucadores como Lazar o David Grusch. Richard Feynman decía que la persona más fácil de engañar era uno mismo. La Ciencia evita que caigamos en nuestras propias trampas mentales. Hasta que no haya pruebas, este tipo de declaraciones no son más que ruido.
¿Debemos investigar los fenómenos anómalos?
Por supuesto. No hay que confundir los reportes de UAP con las declaraciones sobre objetos extraterrestres en oscuras instalaciones militares. Los reportes de fenómenos anómalos están recogidos por instrumentos y radares de aeronaves y son datos objetivos. Otra cosa es si podemos averiguar la causa o no.
¿por qué somos científicos? movidos por la curiosidad, la necesidad de explorar lo que no conocemos, de responder a cuestiones fundamentales que ya nos hacíamos de niños: ¿por qué la hierba es verde, por qué podemos respirar?. Cuando voy a museos o ferias de minerales, siempre hay niños llenos de curiosidad. Pocos niños hay a quienes no les encanten los minerales. Suelen hacer observaciones muy interesantes: ven los aspectos en común que tienen minerales con composición similar, o reconocen las formas cristalográficas. Esa misma curiosidad es la del científico. Lamentablemente, la sociedad tiende a desanimar a esos niños, a destruir su curiosidad. Un científico es alguien que mantuvo esa faceta infantil. De hecho, la curiosidad es el principal, pero el menos valorado activo que posee un científico. Así que, ¿cómo nos resistiríamos a investigar y explorar cualquier fenómeno extraño o anómalo?. No seríamos científicos entonces.
Pero no investigamos de cualquier forma, ni nos dejamos arrastrar por vagos testimonios. Ni afirmamos, ni descartamos nada, sino que planificamos cómo abordar una cuestión o problema. Adoptamos la postura escéptica en el sentido clásico del término, la epojé de Sexto Empírico: suspensión de juicio u opinión durante la observación y experimentación. Por lo que hemos visto anteriormente, es muy poco probable que coincidamos en el tiempo y en la vecindad galáctica con otra forma de vida inteligente. Es poco probable que ésta haya alcanzado un nivel tecnológico suficiente y es poco probable que las propias leyes de la Física permitan tales viajes. Pero no lo descartamos. Simplemente, seguimos trabajando y seguimos las evidencias.
En 2022 la NASA creó una comisión para el estudio de los UAP o ‘fenómenos anómalos no identificados’ (unidentified anomalous phenomena). Este es el nombre correcto, no ‘fenómenos aéreos no identificados’, como indicaron algunos medios, denominación incorrecta que se abandonó.
Estos fenómenos UAP engloban supuestos avistamientos OVNIS (hay cientos de relatos de pilotos en torno a ellos) así como cualquier fenómeno, atmosférico o espacial, no identificado.
La comisión independiente para el estudio de los UAP
Los astrobiólogos debemos estudiar todo fenómeno no explicado o anómalo. No se trata de dar oídos a testimonios, mitos o conspiraciones, sino investigar seriamente qué hay. La mayor parte de los UAP tienen explicaciones sencillas. En muchos casos se han cotejado con tráfico aéreo comercial y se corresponden con detecciones de aviones lejanos.
Aun así, hay fenómenos que, por falta de evidencias o datos, no pueden no ya explicarse, sino ni siquiera estudiarse. Ello no implica que existan inteligencias extraterrestres. Que algo no tenga explicación no implica que sea válida cualquier explicación por extravagante que sea. Por ejemplo, que no pueda explicar el Origen de la Vida no implica que el Origen de la Vida sea obra de Dios. Del mismo modo, que no pueda explicar un borroso objeto que un piloto vio una vez en el aire, no implica que fuera una nave extraterrestre.
La falta de datos, simplemente, implica que, o bien hay que seguir estudiando y obteniendo datos, o, si no podemos tener más datos, quedando la cuestión o hipótesis fuera de toda posibilidad de verificación y de falsación (otra de las herramientas del trabajo científico), la tenemos que dejar archivada. Pero, en la práctica, como decía Sherlock Holmes «en general, cuanto más extravagante es una cosa, menos misteriosa suele resultar»
¿Es posible el mundo de Star Trek?
Como científico debo razonar y usar datos y evidencias. Pero no dejamos de soñar. Desde pequeño, cuando pasaba noches enteras mirando el firmamento, primero con los ojos desnudos, luego con mi pequeño telescopio, siempre soñaba con que alguien más habría en algún planeta ahí arriba. Deseaba que vinieran. Eso implicaría que es posible viajar por el espacio y a otros sistemas planetarios. Si realmente hubiera extraterrestres, ello indicaría que nuestro pequeño mundo podría hacerse mucho más grande. Es una gran esperanza. Pero ello queda fuera del ámbito de la Ciencia.
Que triste sería que no hubiera nadie más en nuestra galaxia. Alguien dijo una vez que somos el Universo contemplándose a sí mismo. Resulta difícil aceptar la idea de que, en 100.000 millones de planetas en nuestra galaxia, no haya otras inteligencias planteándose las mismas preguntas. Pero, más difícil aún es aceptar que, habiendo otras inteligencias, estamos aislados en esta isla planetaria. Así que, como todos los que crecimos con Star Trek (yo lo hice con TNG), soñábamos en cada capítulo que podríamos explorar la galaxia en busca de nuevos mundos y nuevas civilizaciones y que igual, por fin, esa noche se iban a presentar visitantes extraterrestres a resolver nuestras dudas.
Siempre podemos soñar que es posible explorar otros mundos como en Star Trek
Pero, ¿es posible el mundo de Star Trek?. Ello tiene varios problemas: el problema de la probabilidad de vida inteligente, el problema de la tecnología y el problema de la coincidencia de civilizaciones en la misma región del universo que hemos revisado anteriormente.
Pero tiene otro problema: un mundo como el de Star Trek presupone que la evolución no es un fenómeno de caos determinista, sino que es teleológica y que termina en seres humanoides que, incluso, pueden cruzarse formando híbridos. Naturalmente, esta trampa teleológica no tiene sentido desde el punto de vista biológico. En cierto modo, Star Trek implica una profunda aceptación del principio del Diseño Inteligente de origen divino, que culmina en seres a imagen y semejanza de Dios (aunque en Star Trek, en lugar de Dios, se apela a unos seres que sembraron, mediante panspermia de su ADN, los mundos habitables). Dentro de la lógica de Star Trek, es casi un imperativo llegar al conocimiento del viaje interestelar. Pero, desde el punto de vista científico, no hay ninguna razón que sugiera que tales viajes serían tecnológicamente posibles.
Si hay vida en otros planetas, es muy posible que sea similar a nivel molecular, teniendo en cuenta que partimos del mismo espacio químico y las reglas de la química imponen cierto determinismo molecular. Pero, igual que en nuestro planeta, donde el mismo patrón molecular y células similares han dado lugar a innumerables morfologías y posibilidades, no podemos ni imaginar cómo sería una inteligencia extraterrestre. No siquiera sabemos si podríamos identificarla con las herramientas de que disponemos actualmente.
La fauna de Ediacara, que desapareció hace unos 540 millones de años, estaba formada por organismos extraños, como este Hallucigenia. Si la misma biología molecular daba lugar a organismos como éstos, ¿cómo serán los organismos extraterrestres? ¿serán molecularmente similares, aunque morfológicamente inimaginables?
Personalmente, desearía que las declaraciones de David Grusch en el congreso fueran reales. Ojalá haya naves alienígenas, lo que implicaría que es posible encontrarnos y explorar los exoplanetas. Pero no tenemos ni una sola prueba o evidencia. Aquí, tras todo lo dicho, sólo veo dos posibilidades: o nos espera un futuro apasionante o una soledad desoladora.
Conclusiones
La vida es probablemente bastante común en nuestra galaxia, pero la vida inteligente es, probablemente, muy rara o incluso única (nosotros) en este momento.
Por ‘común’ queremos decir en números absolutos. Con lo que conocemos a través de la observación de exoplanetas, la probabilidad de hallar un planeta habitable o con vida es muy baja. Es una situación comparable a la lotería: En todo el país va a tocar el gordo de la lotería seguro en algún sitio, pero la probabilidad de que te toque a tí, en tu barrio o en tu ciudad es baja. Con lo que sabemos por el momento, hay un 99.94% de probabilidad de que un planeta terrestre no tenga vida. Lo que pasa es que la galaxia tiene miles de millones de planetas.
Es extremadamente improbable que exista vida inteligente en nuestro vecindario cósmico o nuestra galaxia, y aún más improbable que dispongan de capacidad para realizar viajes interestelares
Las declaraciones sobre OVNIS y UPA no están respaldadas con ninguna evidencia, solo son rumores. La mayoría de los avistamientos documentados por pilotos y registrados por radares, o bien han sido explicados, o bien no hay datos suficientes como para poder estudiarlos. Los científicos no trabajan con testimonios, sin datos suficientes o incompletos. Esto no quiere decir que no se estudie. Hay que revisar objetivamente todos los datos y documentos.
Aun así, todos nuestros cálculos se basan en la vida terrestre como modelo. Esto podría cambiar, por lo que seguimos estudiando e investigando y nos mantenemos alertas.
Referencias
Ćirković, M. M. (2013) ‘Who are the SETI sceptics?’, Acta Astronautica, 89, pp. 38–45. doi: 10.1016/j.actaastro.2013.03.012.
Dick, S. J. (2012) ‘Critical Issues in the History, Philosophy, and Sociology of Astrobiology’, Astrobiology, 12(10), pp. 906–927. doi: 10.1089/ast.2011.0786.
Haqq-Misra, J., Kopparapu, R. K. & Schwieterman, E. (2020) ‘Observational Constraints on the Great Filter’, Astrobiology, 20(5), pp. 572–579. doi: 10.1089/ast.2019.2154.
Kipping, D. (2020) ‘An objective Bayesian analysis of life’s early start and our late arrival’, Proceedings of the National Academy of Sciences, 117(22), pp. 11995–12003. doi: 10.1073/pnas.1921655117.
Schmidt, G. A. and Frank, A. (2019) ‘The Silurian hypothesis: would it be possible to detect an industrial civilization in the geological record?’, International Journal of Astrobiology, 18(2), pp. 142–150. doi: 10.1017/S1473550418000095.
Seckbach, J. (2012) Genesis – In The Beginning, Cellular Origin, Life in Extreme Habitats and Astrobiology, Vol. 22. Edited by J. Seckbach. Dordrecht: Springer Netherlands (Cellular Origin, Life in Extreme Habitats and Astrobiology). doi: 10.1007/978-94-007-2941-4.
Spada, G. and Melini, D. (2020) ‘Evolution of the number of communicative civilizations in the Galaxy: implications on Fermi paradox’, International Journal of Astrobiology, 19(4), pp. 314–319. doi: 10.1017/S1473550420000063.
Webb, S. (2015) If the Universe Is Teeming with Aliens … WHERE IS EVERYBODY?, If the Universe is Teeming with Aliens… Where is Everybody? Cham: Springer International Publishing (Science and Fiction). doi: 10.1007/978-3-319-13236-5.
HEXOKINASA: ESTRUCTURA, EVOLUCIÓN Y PAPEL EN EL CÁNCER
escrito por cmfc5_1A | 17 diciembre, 2024
Redactado por María Arrondo Sánchez y Carolina Amil Zamorano
INTRODUCCIÓN
La hexoquinasa es una enzima transferasa (D-hexose-6-phosphotransferase), del grupo de las quinasas, encargada de fosforilar hexosas. Sin embargo, presenta mayor afinidad por la glucosa, puesto que la Km de esta es menor que la de otras hexosas como la fructosa. Esta proteína presenta cuatro isoformas, que han ido surgiendo de forma gradual. En dicho proceso de evolución ocurren determinados cambios que son claves en la estructura y que han permitido que la hexokinasa en dos de sus isoformas se oligomerice. La estructura de la proteína va a contar con dos dominios de unión a los sustratos (glucosa y ATP) y su actividad va a estar regulada alostéricamente mediante un mecanismo de ajuste inducido provocado por la propia glucosa.
Así mismo, esta enzima va a presentar un papel clave en el cáncer. En este artículo se abordará el normal funcionamiento de la hexokinasa así como su papel tumoral por diferentes vías, profundizando, además, en posibles estudios futuros y nuevos campos que se abren en la investigación contra el cáncer que emiten un rayo de esperanza en el estudio biomédico.
PAPEL BIOLÓGICO
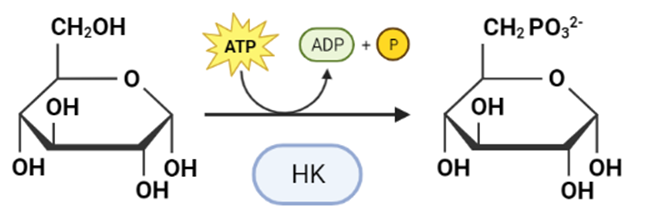
Figura I: reacción esquemática de la Hexokinasa, que fosforila la glucosa, produciendo Glucosa-6-Fosfato.
La Hexokinasa participa en la primera reacción irreversible de la glucólisis, que es la primera etapa del metabolismo de la glucosa. Tras hidrolizar el ATP, transfiere el grupo fosfato a la glucosa, para dar glucosa-6-fosfato (G6P), y así manteniendo el gradiente de glucosa que permite que haya un flujo de la misma mediado por los transportadores GLUT. La G6P es un inhibidor competitivo del ATP, por tanto se trata de un fenómeno de feedback negativo, en el que el mismo producto regula alostérica y negativamente su reacción de síntesis. Además, el grupo fosfato (Pi) liberado de la hidrólisis del ATP puede antagonizar la inhibición de G6P o sumarse al efecto inhibidor, según la isoenzima que haya llevado a cabo la reacción.
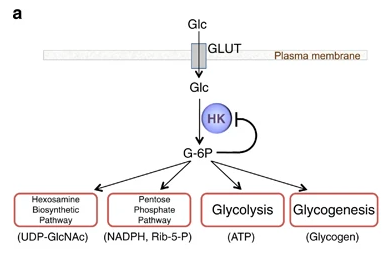
El producto (G6P) puede seguir varias rutas o vías celulares y funcionales:
Metabolismo catabólico: se introduce la glucosa en la glucólisis, para llevar a cabo un metabolismo oxidativo y obtener energía.
Metabolismo anabólico: la G6P es destinada a la vía de las pentosas fosfato, para sintetizar NAPDH y Ribulosa-5-Fosfato; o puede ser convertido a sus formas poliméricas (glucógeno), mediante la gluconeogénesis.
Figura II: vías del producto Glucosa-6-Fosfato, que se introduce en vías como la glucólisis, la ruta de las pentosas fosfato, o la glucogénesis.
ESTRUCTURA
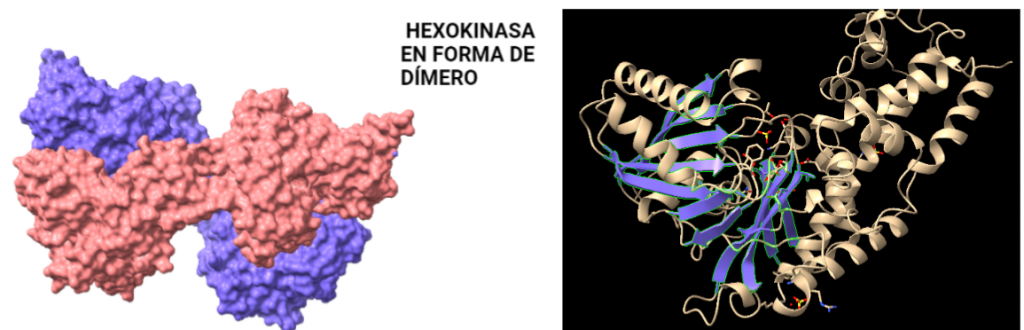
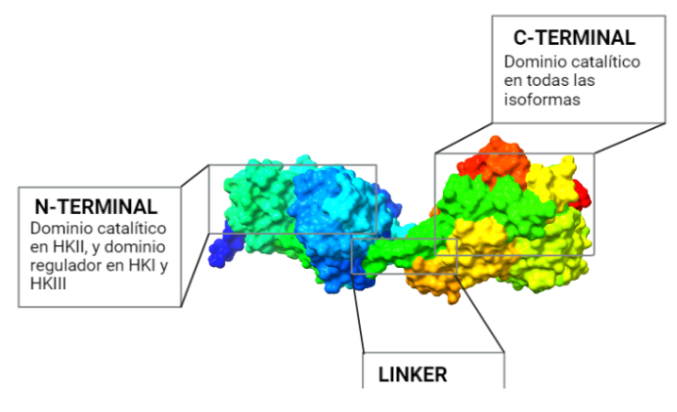
La estructura de las Hexokinasas más comunes (las isoformas I, II y III) cuenta con dos lóbulos muy similares de unos 50KDa cada uno. Algunas de ellas, como la HK I, son monoméricas, pero cuando se une a la membrana externa de la mitocondria se oligomerizan. De esta manera, la Hexokinasa cuenta con dos dominios principales, uno regulador y otro catalítico. La estructura dimérica y por tanto cuaternaria está presente en todas las isoformas salvo en la IV, que es la más ancestral.
Figura III: HexokinasaI dimérica, con cada monómero de un color. Hecho con BioRender y Chimera, a partir de PDB 1BG3. Figura IV: Dominio de unión del ATP, se observan cuatro láminas paralelas y una antiparalela. Hecho con Chimera
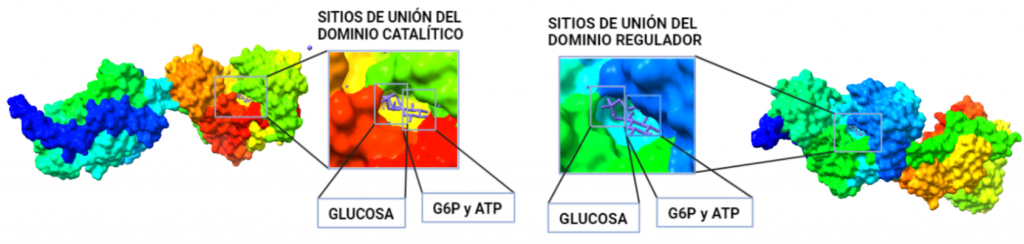
El dominio N-terminal se considera el dominio regulador en las isoenzimas I y II, y contiene el motivo de unión a la mitocondria. Además, está unido al dominio C-terminal (que es el dominio catalítico) a través de una hélice alfa. Ambos dominios presentan sitios de unión con la glucosa, G6P y ATP, y la inhibición de G6P en el dominio regulador se contagia al dominio catalítico por medio del contacto por la hélice alfa entre los dominios. La estructura terciaria de la hexoquinasa se basa en un plegamiento alfa/beta abierto. El dominio de unión al ATP está compuesto por cinco láminas beta y tres hélices alfa en el cual cuatro de las láminas beta son paralelas y una es antiparalela. Por otro lado, la hexoquinasa requiere de iones de magnesio para poder llevar a cabo la actividad catalítica. El magnesio (Mg2+) va a ser el cofactor de la enzima y se encuentra formando un complejo con el ATP (MgATP2-), que estabiliza la catálisis y reduce la energía de activación de la reacción.
Figura V: esquema general de la Hexokinasa en forma de monómero, que presenta un dominio catalítico y otro regulador, unidos por una hélice alfa. Hecho con BioRender, a partir de PDB 1BG3.Figura VI: Sitios de unión de la Hexokinasa con la glucosa y el inhibidor G6P, en ambos dominios. Hecho con BioRender, a partir de PDB 1BG3.
EVOLUCIÓN
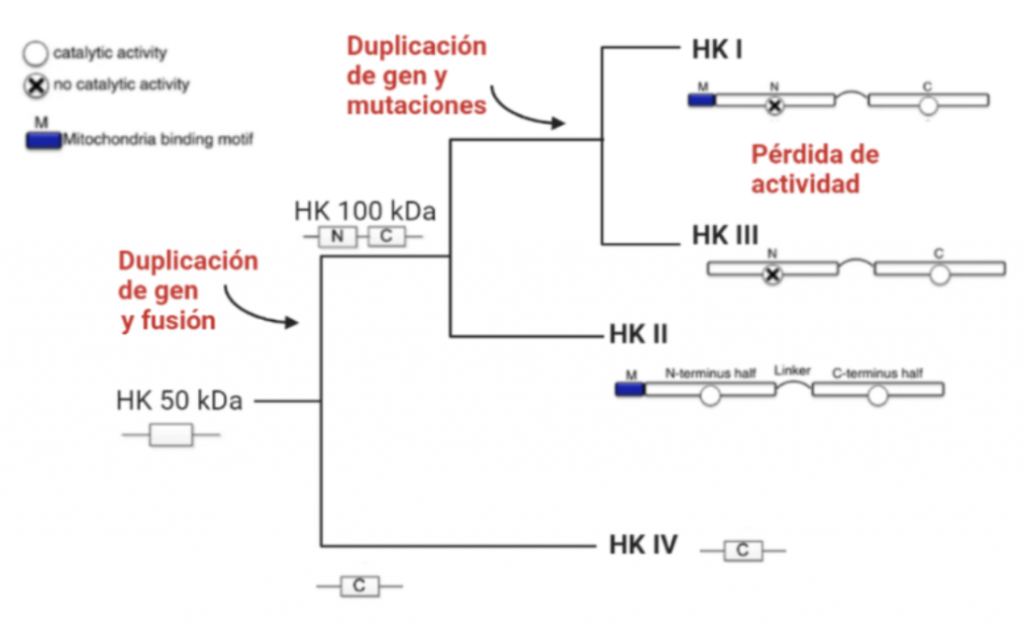
Todas las isoenzimas de la Hexokinasa provienen de una Hexokinasa de 50 kDa, susceptible a la inhibición por el producto G6P, por tanto, todas las isoenzimas presentan esta característica. A partir de la duplicación y fusión del gen que codificaba esta forma ancestral, surgieron las isoenzimas I, II y III, que ya son moléculas de 100kDa.
Figura VII: Esquema de la evolución de las isoformas de la Hexokinasa, hecho con BioRender e imágenes de D J Roberts y S. Miyamoto
La isoforma más próxima evolutivamente a la Hexokinasa original es la Tipo IV, que no sufrió la duplicación y fusión génica. Una vez que esto ocurrió, la segunda isoenzima que apareció fue la Hexokinasa II, que mantiene la actividad catalítica en ambos extremos terminales de la proteína, al igual que la Hexokinasa ancestral.
Una consecuente duplicación tuvo como resultado la aparición de la isoforma III. Posteriormente, las mutaciones de genes que codificaban la Hexokinasa 100 kDa, produjeron que el extremo N-terminal se diferenciara funcionalmente, perdiendo la actividad catalítica, y adquiriendo una función reguladora (con un sitio de unión para el inhibidor G6P). Esta diferenciación dio lugar a las en las Hexokinasas I y III.
Además, en las HK I y II, el extremo N-terminal presenta un dominio hidrofóbico que permite a estas integrarse en la membrana de la mitocondria. Concretamente, se unen a las porinas (VDAC) de la membrana mitocondrial externa, las cuales interaccionan con los ANT (Translocadores de Nucleótidos de Adenina). Esto es esencial para el mecanismo enzimático de la HK, puesto que es el sitio de salida del ATP producto de la fosforilación oxidativa (que usará la HK), y el sitio de entrada del ADP resultante de la reacción enzimática de la hexokinasa. Por tanto, existe una coordinación entre la introducción de la glucosa al metabolismo glucolítico y las últimas etapas de este en la mitocondria (la fosforilación oxidativa), para que se den a un ritmo adecuado a las necesidades celulares.
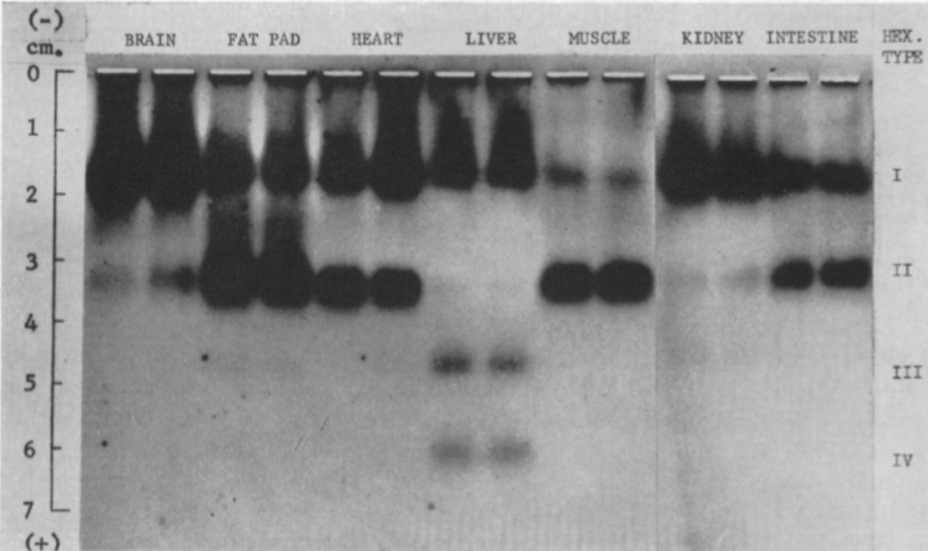
Hay cuatro isoenzimas de la Hexokinasa (HK) en los tejidos de mamíferos, con una estructura similar, pero expresión en diferentes tejidos:
Figura VIII: Electroforesis de las isoformas de la Hexokinasa en diferentes tejidos, que muestra que la HK I se encuentra presente de manera general, mientras que la HK II aparece en el músculo y tejido adiposo, y la HK III y IV, en el hígado. Imagen de H M Katzen and R T Schimk.
HEXOKINASA I (HKI)
Fundamentalmente en el cerebro, donde la tasa metabólica es muy exigente, pero expresada de manera general.
HEXOKINASA II (HKII)
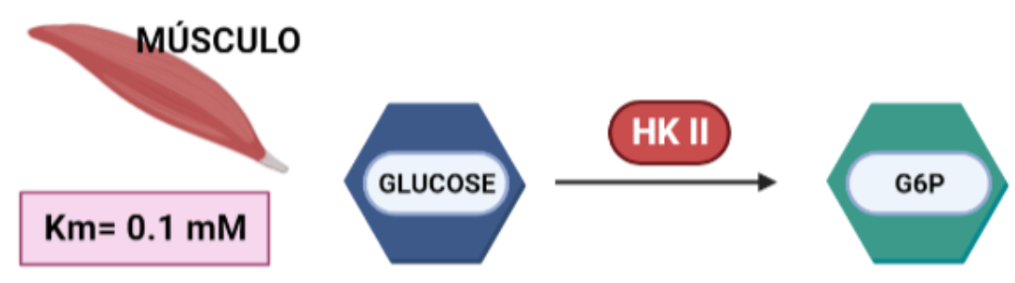
Más limitada en su expresión, aparece en tejidos sensibles a insulina, como es el caso del tejido adiposo y el músculo esquelético. En el músculo, es necesaria una alta tasa de glucólisis, y por tanto, lo que ocurre es que esta isoforma presenta una gran afinidad por la glucosa (menor Km), para que con concentraciones muy bajas de glucosa, se alcancen altas velocidades de la enzima.
Figura IX: Afinidad por la glucosa de la HKII en músculo. Creado con BioRender
La insulina aumenta la actividad de esta isoforma, induciendo la transcripción del gen que la codifica, y de esta forma, favoreciendo la metabolización y eliminación de glucosa. Por esta razón, en los individuos con diabetes de tipo II, la expresión de la HKII se ve reducida, acentuando la hiperglucemia.
Al igual que la isoenzima Tipo I, incluye un dominio hidrofóbico en el extremo N-terminal que permite que se inserte en la membrana externa mitocondrial, y también usa ATP intramitocondrial.
Cabe destacar su predominancia en células tumorales, puesto que estas se caracterizan por presentar un aumento anormal del metabolismo, en el que la reacción que lleva a cabo la Hexokinasa es esencial para la obtención de energía.
HEXOKINASA III (HKIII)
A diferencia de las dos isoenzimas anteriores, la Hexokinasa III no está unida a la mitocondria, puesto que carece del dominio hidrofóbico en el extremo N-terminal. Se piensa que se expresa en el citoplasma, o que incluso tiene una localización perinuclear, en células del hígado.
HEXOKINASA IV (HK IV)
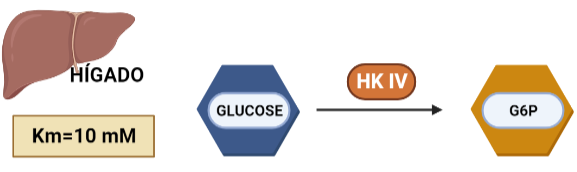
En hepatocitos y células beta pancreáticas, y es conocida como glucoquinasa. La G6P que produce está destinada a la síntesis deglucosa. Esto es un proceso que se lleva a cabo cuando la cantidad de sustrato (glucosa) es alta, por tanto, tiene sentido que esta isoenzima presente una mayor Km, porque necesitará altas concentraciones de glucosa para realizar la reacción a una velocidad alta.
Figura X: Afinidad por la glucosa de la HK IV en hígado. Creado con BioRender.
MECANISMO
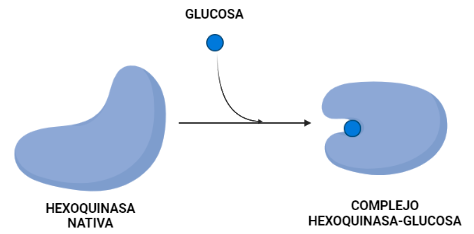
La hexoquinasa sufre un cambio conformacional que es regulado por la propia glucosa que va a ser esencial para la catálisis. En este proceso se observa que la superficie en contacto con el solvente del complejo hexoquinasa-glucosa es más pequeña que la hexoquinasa nativa.
Utilizando dicho cambio en el área de superficie que se encuentra expuesta se ha podido estimar la contribución hidrofóbica a los cambios de energía libre tras la unión de la glucosa. De esta manera se descubre que el efecto hidrofóbico por sí solo favorece la conformación activa de la hexoquinasa en presencia y ausencia de azúcar. La estabilidad observada de la conformación inactiva de la enzima en ausencia de sustratos puede resultar de una deficiencia de interacciones complementarias dentro de la cavidad que se forma cuando los dos lóbulos se unen.
El cambio conformacional que sufre la hexoquinasa mantiene la estructura terciaria prácticamente igual excepto por un gran cambio en la orientación de los dos lóbulos. Para demostrar este cambio lo que se hizo fue superponer los carbonos alfa de cada lóbulo usando un procedimiento de mínimos cuadrados y tratando a los carbonos como cuerpos rígidos. Esta superposición mostró que cada lóbulo se comporta como un cuerpo rígido durante el cambio conformacional entre la forma nativa de la proteína y el complejo con la glucosa.
Figura XI: Cambio conformacional de la HK inducido por la glucosa y cambio en la superficie de contacto con el solvente.
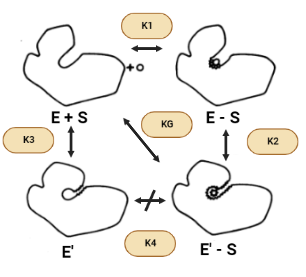
Tal y como venimos viendo, la superficie accesible de la hexoquinasa se reduce cuando se une la glucosa para formar el complejo enzima-sustrato (ES) y se reduce aún más por el cambio a E’(inactiva)-S. Por lo tanto, se puede esperar que las fuerzas hidrofóbicas favorezcan la conformación activa en presencia de azúcar, asumiendo que todos los donantes y aceptores de enlaces de hidrógeno están satisfechos en E’-S. Sin embargo, el área superficial también se reduce cuando la conformación activa se forma en ausencia de azúcar (E’ – E). Sin embargo, debido a que hay menos de un factor de dos diferencias entre los cambios en la superficie accesible para las transiciones E-S → E’-S y E’-E, el efecto hidrofóbico no puede explicar la gran diferencia en las constantes de equilibrio conformacional K2 y K3 en presencia y ausencia de azúcar.
Figura XII: Cambios conformacionales de la Hexoquinasa en presencia y ausencia de azúcar. Esquema creado con Biorender.
Otra cuestión que surge es por qué la enzima no permanece en el estado activo, E, en ausencia de ligandos sabiendo que el efecto hidrófobo, de manera individual, predice que la estructura E debería ser más estable. La respuesta a esta pregunta reside en que cuando la enzima carece de la presencia de la glucosa contiene una cavidad en la que entran las moléculas de agua y donde quedan encerradas. Además, tanto los puentes de hidrógeno como las fuerzas de Wan der Waals contribuyen muy poco a la estabilidad de la proteína y del complejo proteína-ligando. El hecho de no obtener estas interacciones complementarias dentro de la cavidad daría como resultado entalpías desfavorables causadas por la pérdida de los puentes de hidrógeno o fuerzas de Van der Waals en relación con los que se producen en la estructura abierta. También puede haber alguna pérdida de entropía traslacional al atrapar una pequeña cantidad de moléculas de agua en la cavidad.
Presuntamente, el agua misma desestabiliza la forma activa mediante la creación de interacciones favorables con la estructura abierta inactiva. Solamente el ligando correcto puede proporcionar las fuerzas de Van der Waals y los puentes de hidrógeno necesarios para que se active la estructura.
Con ello, concluimos que hay al menos dos posibles funciones para el cambio conformacional inducido por la glucosa: permitir un «mecanismo de acogida» o proporcionar especificidad.
PAPEL BIOMÉDICO: HEXOKINASA II EN CÁNCER
El metabolismo de las células tumorales se caracteriza por una alta actividad glucolítica: metabolizan anaeróbicamente grandes cantidades de glucosa en ácido láctico, incluso en presencia de oxígeno, aumentando la velocidad de la glucólisis y de la síntesis de ATP. Esto es lo que se conoce como el efecto Warburg. Por tanto, la actividad de cualquier enzima glucolítica como la HK será esencial en un tumor.
La Hexokinasa II aparece sobreexpresada en células tumorales, satisfaciendo estas altas velocidades de la glucólisis. La introducción de la glucosa en el metabolismo glucolítico es crucial para la producción de energía, y la síntesis de precursores de nucleótidos (derivados de glucosa) por la vía de las pentosas fosfato, destinados a la síntesis de ADN para la proliferación del tumor.
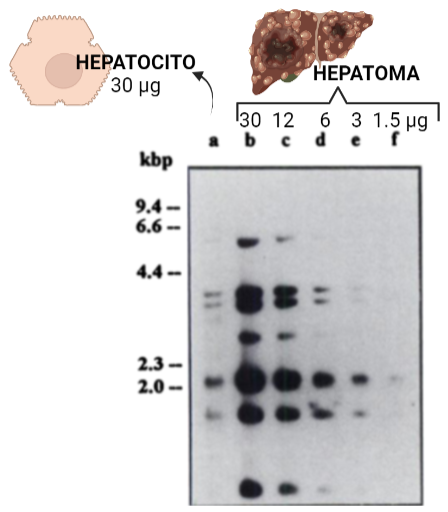
Fig. XIII: Amplificación del gen de la HKII en hepatoma y hepatocitos. Hay unas 5-10 copias más de lo normal en el hepatoma, ya que la intensidad de una tira de ADN de hepatocito de 30 μg se asemeja a la de ADN de hepatoma de 6-3 μg Imagen: Annette Rempel
La regulación por AKT de la HKII es el factor determinante para que esta isoforma sea la esencial en el metabolismo tumoral, puesto que controla la unión de esta a la mitocondria, y con ello, fija la función de la HK, que varía en función de si aparece unida a la mitocondria o no.
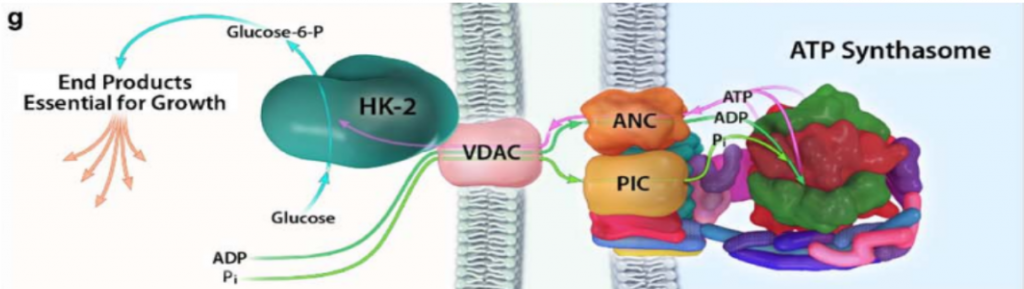
Con el objetivo de crecimiento del tumor, y cuando hay disponibilidad de nutrientes, AKT une la HKII a la mitocondria (fosforilando su residuo Thr-473), conectándola con VDAC. Esto le permite a la enzima tener un acceso privilegiado al ATP que sintetiza la mitocondria, y la célula sigue un metabolismo de proliferación y de producción de energía, mediante la glucólisis.
Figura XIV: Unión de la HKII con el VDAC de la membrana externa de la mitocondria, y cómo esta le posibilita acceder al ATP sintetizado por la ATP sintasa. Esta imagen señala la relación entre el primer paso de la glucólisis y el último paso del metabolismo oxidativo. Imagen: Pedersen, P.L.
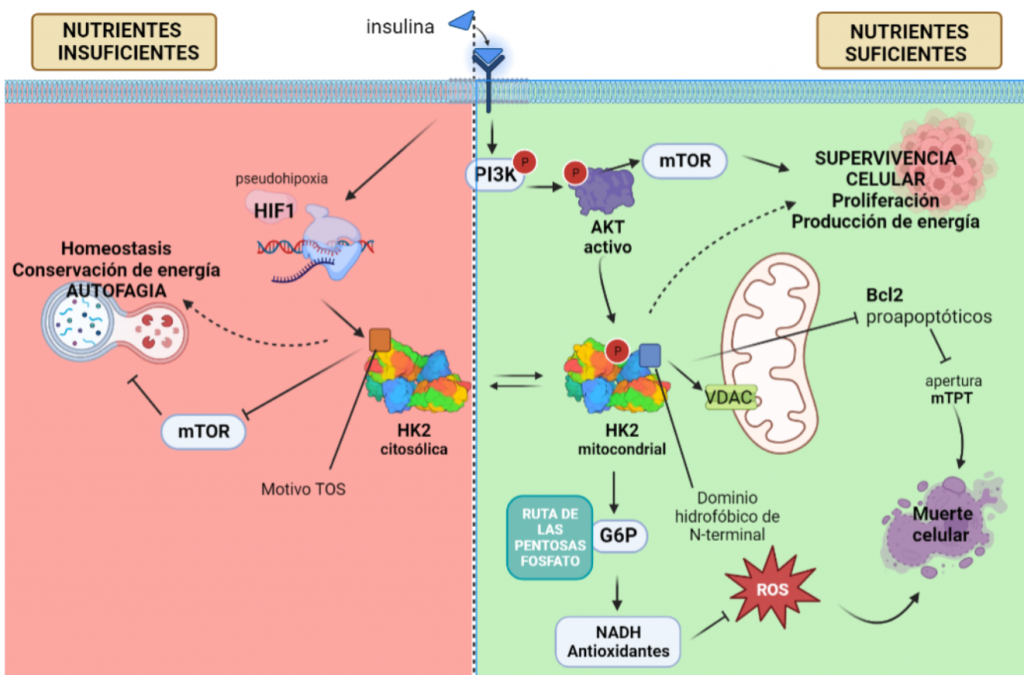
La HK II mitocondrial, además, lleva a cabo una función protectora en tumores con acceso a nutrientes, promoviendo la supervivencia celular: inhibe a los miembros pro-apoptóticos de la familia Bcl2, e introduce la G6P en la ruta de las pentosas fosfato, cuyos productos son antioxidantes que reducen las ROS (especies reactivas de oxígeno).
Sin embargo, en una situación de isquemia, en la que no llegan suficientes nutrientes y oxígeno a la célula, disminuye la actividad de AKT y HKII mitocondrial, aumentando la HKII citoplásmica. Entonces, esta isoforma se une a mTORC (mediante el motivo TOS), inhibiéndola, e induciendo la vía autofágica, y la conservación de energía y homeostasis en ausencia de glucosa, pensando en el “bien mayor” del tumor.
Figura XV: Esquema explicativo del papel que juega la HK II en células tumorales, promoviendo la supervivencia celular si tiene acceso a nutrientes, pero induciendo indirectamente la autofagia en el caso contrario, con el fin de conservar la energía y homeostasis tisular.Creado con BioRender.
Esta regulación por AKT posibilita esta compleja acción de la HKII, que puede ser proapoptótica (HKII citoplásmica) o antiapoptótica (HKII mitocondrial), según la disponibilidad de recursos. Sin embargo, AKT no regula la HKI, ya que esta no presenta una secuencia de consenso para esta enzima, y por ello, esta isoenzima no se encuentra prevalentemente en tumores. Además, la HKI no puede satisfacer la alta demanda energética, al perder la actividad catalítica en el extremo N-terminal.
Según todas las vías beneficiosas para el tumor mencionadas anteriormente en las que participa la HKII, la eliminación de esta isoforma perjudicaría a la progresión del tumor, por lo tanto, es un frente esperanzador en terapias oncogénicas. Lo ideal sería encontrar un modo de inhibir únicamente esta isoforma, pero es difícil puesto que todas ellas son bastante similares.
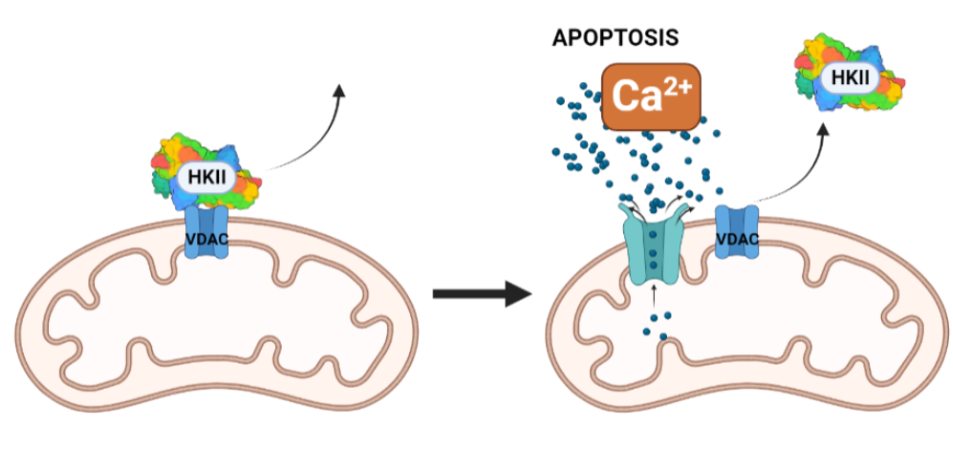
La inhibición de la HKII en células cancerosas puede darse por p53, o por un sustrato análogo a la glucosa, la 2-desoxiglucosa, que favorece la apoptosis. Una combinación de estos dos factores podría ser favorecedora. Otro posible campo a investigar sería la inhibición de la HKII por fosfato inorgánico, que sensibiliza la inhibición por G6P en esta, pero la antagoniza en HKI. Incluso, una alternativa más podría ser el uso de determinados péptidos que desplazaran la HKII de la mitocondria. Este desplazamiento parece producir un aumento de la concentración de Ca2+ citosólica, lo que abriría poros en la membrana mitocondrial e induciría a la célula a apoptosis.
Figura XVI: Posible terapia oncogénica sobre HK II, mediante péptidos que la separen de la mitocondria. Creado con BioRender.
Bennett, W. S., & Steitz, T. A. (1978). Glucose-induced conformational change in yeast hexokinase. Proceedings of the National Academy of Sciences, 75(10), 4848-4852. https://www.pnas.org/doi/abs/10.1073/pnas.75.10.4848
Gahr, M. (1980). Isoelectric Focusing of Hexokinase and Glucose-6-Phosphate Dehydrogenase Isoenzymes in Erythrocytes of Newborn Infants and Adults. British Journal of Haematology, 46(4), 529-535. https://doi.org/10.1111/j.1365-2141.1980.tb06009.x
Haruhiko Osawa, Calum Sutherland, R. Brooks Robey, Richard L. Printz, Daryl K. Granner (1996). Analysis of the Signaling Pathway Involved in the Regulation of Hexokinase II Gene Transcription by Insulin. https://www.sciencedirect.com/science/article/pii/S002192581831932X
Mulichak, A., Wilson, J., Padmanabhan, K. et a (1998)l. The structure of mammalian hexokinase-1. Nat Struct Mol Biol 5, 555–560. https://pubmed.ncbi.nlm.nih.gov/9665168/
R.L. Printz, S. Koch, L.R. Potter, R.M. O’Doherty, J.J. Tiesinga, S. Moritz, D.K. Granner (1993). Hexokinase II mRNA and gene structure, regulation by insulin, and evolution. Journal of Biological Chemistry. Volume 268, Issue 7 https://www.sciencedirect.com/science/article/pii/S0021925818535213
Reference for PDB-101: PDB-101: Educational resources supporting molecular explorations through biology and medicine. Christine Zardecki, Shuchismita Dutta, David S. Goodsell, Robert Lowe, Maria Voigt, Stephen K. Burley. (2022) Protein Science 31: 129-140 doi:10.1002/pro.4200
Reference for RCSB PDB: The Protein Data Bank H.M. Berman, J. Westbrook, Z. Feng, G. Gilliland, T.N. Bhat, H. Weissig, I.N. Shindyalov, P.E. Bourne (2000) Nucleic Acids Research, 28: 235-242. doi:10.1093/nar/28.1.235
Rempel, A., Mathupala, S. P., Griffin, C. A., Hawkins, A. L., & Pedersen, P. L. (1996). Glucose Catabolism in Cancer Cells: Amplification of the Gene Encoding Type II Hexokinase1. American Association for Cancer Research. https://pubmed.ncbi.nlm.nih.gov/11557773/
Roberts, D., Miyamoto, S. (2015) Hexokinase II integrates energy metabolism and cellular protection: Akting on mitochondria and TORCing to autophagy. Cell Death Differ 22, 248–257. https://www.nature.com/articles/cdd2014173
UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Goddard TD, Huang CC, Meng EC, Pettersen EF, Couch GS, Morris JH, Ferrin TE. Protein Sci. 2018 Jan;27(1):14-25.
UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Pettersen EF, Goddard TD, Huang CC, Meng EC, Couch GS, Croll TI, Morris JH, Ferrin TE. Protein Sci. 2021 Jan;30(1):70-82.